



Dietary fat and its influence on the rumen microbiota.
Ruminant digestive processes, especially those like carbohydrate fermentation and dietary protein degradation, are carried by rumen microbiota, which includes: bacteria, protozoa, fungi and archaea.
This symbiotic microbiota provides nutrients to the host. Amongst which the most significant are: volatile fatty acids (VFA), microbial protein, and vitamins. However, within the process of microbial fermentation, energy losses due to methane production are also very present.
Despite high rumen stability due to its functional reliability and its microbial ecosystem resilience (Weimer et al, 2015), rumen microbiota presents large individual variations (Jami and Mizrahi 2012) and can be altered by abrupt or significant dietary changes. An example of this are dietary changes in starch or fat content.
Fatty supplements began to be used as a means to increase dietary energy values and meet energy requirements in dairy cows and fattening cattle within intensive production systems. This type of supplements can also be used to modify fatty acid (FA) profiles of meat (Wood et al., 2008) and milk, modulating its dietary, organoleptic and technological properties.
In most ruminant diets, fat accounts for less than 5% of total dry matter. Having oilseeds like flaxseeds, rapeseed, soybeans and sunflower as the main sources of fat for dietary inclusion. These seeds are rich in unsaturated fatty acids (UFA).
- Oleic acid (OA, cis-9-C18:1)
- Linoleic acid (LA)
- Alpha-linolenic acid (ALA)
1st step. Lipolysis
Lipolysis is the first step for ruminal metabolism of acylglycerols. Resulting in the release of FAs.
Adding fat to the diet can:
+ Modulate rumen function, thus reducing methane emissions (Martin et al., 2016)
– Decrease intake and fat content in milk (Rabiee et al., 2012)
A limiting factor of fat supplementation in ruminant diets, is the negative effect that they exert on rumen degradability. This is especially true when there is a high proportion of UFAs (Brooks et al., 1954).
This entry focuses on the influence of fat on rumen microbiota.
Most FAs found in ruminant diets are glycerol esters:
- TRIACYLGLYCEROLS, mainly in concentrates
- GALACTOLIPIDS AND PHOSPHOLIPIDS mainly found in in forage. Except in silages where plant lipases release fatty acids.
Figure 1
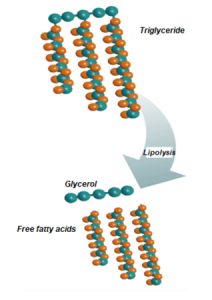

Lipolysis: It refers to the action of lipases on the ester bonds that bind fatty acids and alcohols. When these are hydrolyzed, fatty acids are released into the medium(Scheme 1). This process is carried out by microbial lipases, although plant material also holds some lipolytic activity.
2º step . Detoxifying adaptation – biohydrogenation
Rumen microbiota completely saturates UFAs.

This process is considered as a detoxifying adaptation (Kemp et al., 1984), and marginally contributes to the elimination of reducing equivalents produced by rumen fermentation (Lourenço, et al. 2010).
Protozoa encompass bacteria, and bacterial biohydrogenation can take place within protozoa (Jenkins et al., 2008) which explain their high concentration of intermediate products (Devillard et al., 2006).
Kim et al. (2002) isolated a bacterium identified as Megasphaera elsdenii YJ-4 that produces trans-10, cis-12-CLA in the rumen under a starch diet and further found that the T81 strain also produces this isomer.
Two bacteria belonging to the genus Fusocillus were also shown to reduce C18:1 FA to stearic acid (Harfoot 1978).
Van de Vossenberg and Joblin (2003) isolated a strain of Butyrivibrio capable of completing the BH of both AL and ALA to stearic acid.


Wallace (2007) concluded that stearic acid-forming bacteria previously identified as Fusocillus sp. or Bacillus hungatei occupy a specific branch of the Butyrivibrio tree.
Biohydrogenation (BH) comprises several steps, depending on the UFAs, and several pathways, depending on the diet and rumen environment (Griinari et al., 1998).
- Phospholipids and galactolipids can be hydrolyzed by some strains of Bacillus fibrisolvens (Hazlewood and Dawson 1979).
- Triacylglycerols are also hydrolyzed by different species of the Butyrivibrio group (Latham et al., 1972), but Anaereovibrio lipolyticus is the best-known triglyceride hydrolyzing bacterium.
- Anaereovibrio lipolyticus lipase was first studied by Henderson (1971) and its genome contains three lipase encoding genes(Prive et al., 2013).
- All three enzymes were more active against lauric and myristic than against palmitic or stearic, while dietary fats contain mainly 16- and 18-carbon FAs.
In vitro studies
Beyond the studies based on the selected isolates, in vivo and in vitro trials have been carried out to evaluate the relationship between rumen bacteria and biohydrogenation (BH). This has been done by adding bacteria and quantifying their products, or by adding dietary supplements known to affect BH and measuring bacterial abundance.
Inoculating B. fibrisolvens in the rumen of goats fed an oil-enriched diet high in linoleic acid (AL) increased concentrations of total conjugated linoleic acid (CLA) within ruminal fluid. Confirming that this bacterium is involved in in vivo biohydrogenation (Shivani et al., 2016).
Due to the fact that most dietary fats are glycerides and need to undergo lipolysis before BH, a reduction in FA metabolism within the rumen could limit ruminal BH.
A preliminary study with esterase inhibitors has shown promising in vitro results(Sargolzehi et al., 2015).
Apás et al. (2015) showed a higher proportion of cis-9, trans-11-CLA in goat milk supplemented with a mixture of Lactobacillus, Bifidobacterium and Enterococcus.


Dietary fat shapes rumen microbiota. Brooks et al (1954) showed that corn oil inclusion both in vitro and in vivo decreases the degradation of rumen cellulose and the production of VFAs by affecting the microbiota. They also found that lard, which has greater saturation compared to corn oil, produces a smaller reduction in cellulose degradation.
Similarly, Ikwuegbu and Sutton (1982) found that the inclusion of flaxseed oil, generated a reduction in fiber degradability, acetate and butyrate percentage, and an increase in propionate .
Plant extracts could also modulate the activity of bio-hydrogenating bacteria.
- Essential oils decreased or increased (Ishlak et al., 2015) the abundance of fibrisolvens in vitro, which could explain changes in the profile of BH products in the rumen (Lourenço, et al. 2010).
- Similarly, tannins decreased the abundance of proteoclasticus and increased B. fibrisolvens (Ishlak et al., 2015) in vitro.


It should be noted that most of these experiments were conducted with oils, and that most of the experimental amounts exceed those used in the field.
Accordingly, experimental results regarding the effects of added fat on the microbiota and its activity should be analyzed with caution, before extrapolating such findings to field conditions.
In the second installment, the obtained results will be analyzed for the in vivo trials. As well as looking a little deeper into fat biohydrogenation in the rumen. Which as previously mentioned, is the step that follows lipolysis when fats have entered the rumen.