


De fato, os ácidos orgânicos associados com específica atividade antimicrobiana, são os ácidos de cadeia curta (C1 – C7) que são ácidos monocarboxílicos simples, como ácidos fórmico, acético, propiônico e butírico, ou são ácidos carboxílicos contendo um grupo hidroxila, como láctico, málico, tartárico e cítrico (Ricke, 2003).
→ Produzem acidez, a qual por sua vez age como flavorizante e também retarda a degradação enzimática.
→ Atuam como agentes quelantes que se ligam a metais formando os quelatos metálicos, os quais previnem/reduzem a oxidação oriunda da catálise dos metais-íons.
→ Agem diretamente como fortes inibidores do crescimento microbiano podendo ter uso na preservação de grãos e rações, sanitização da carne e como aditivo promotor de crescimento na ração.
pH do trato digestivo, capacidade tampão dos ingredientes da dieta, presença de outros antimicrobianos na dieta, condição higiênica do ambiente produtivo e heterogeneidade da flora intestinal (Dibner e Buttin, 2002; Ricke, 2003).


Alguns ácidos orgânicos, particularmente os ácidos graxos de cadeia curta (AGCC), acetato, propionato e butirato, são produzidos em quantidade milimolar no trato gastrointestinal dos animais e, caracteristicamente, ocorrem em altas concentrações em regiões onde há microbiota estritamente anaeróbica predominante.
→ Como os antibióticos, os ácidos orgânicos de cadeia curta têm uma atividade antimicrobiana específica e, ao contrário dos antibióticos, esta atividade antimicrobiana dos ácidos orgânicos depende do pH do meio.
Reduções no número de bactérias estão associadas à alimentação com ácidos orgânicos, que são particularmente eficazes contra espécies de bactérias intolerantes aos ácidos, como E. coli, Salmonella e Campylobacter (Dibner, 2002).
Essa redução da concorrência é um dos mecanismos responsáveis por uma melhor digestibilidade, a qual foi relatada em suínos por numerosos pesquisadores (Partanen, K. 2001; Eidelsburger et al., 1992; Mroz et al. 1997; Mroz et al., 2000; Blanck et al., 1999; Kemme et al., 1999; Jongbloed et al., 2000.), sendo que essas melhorias na digestibilidade foram associadas a melhorias significativas no desempenho dos animais.
Os ácidos orgânicos têm vários efeitos adicionais, que vão além dos antibióticos. Isso inclui
→ redução no pH da digesta
→ aumento da secreção pancreática
→ efeitos tróficos na mucosa gastrointestinal.


Em geral, possíveis alvos dos ácidos orgânicos incluem a parede celular, membrana citoplasmática e funções metabólicas específicas no citoplasma, associadas com a replicação e síntese de proteínas (Denyer e Stewart, 1998; Davidson, 2001).
Os numerosos relatórios de atividade antimicrobiana in vitro e in vivo deixam pouca dúvida de que os ácidos orgânicos exercem parte de seus efeitos através da redução de populações microbianas gastrointestinais, particularmente entre espécies sensíveis ao ácido (Dibner e tal., 2002).
A importância do pH baixo nas atividades antimicrobianas dos ácidos orgânicos, pode ser explicada pelo seu efeito na dissociação do ácido.
Em pH baixo, mais ácido orgânico estará na forma não dissociada. Ácidos orgânicos não dissociados são lipofílicos e podem se difundir através das membranas celulares de bactérias e fungos (Partanen, 2001).
→ As formas não dissociadas de ácidos orgânicos podem, facilmente, penetrar na membrana lipídica da célula bacteriana e, uma vez no interior onde o pH do citoplasma celular é neutro, o ácido orgânico se dissocia em ânions e prótons (Eklund, 1983, 1985; Salmond et ai., 1984; Cherrington et al., 1990, 1991; Davidson, 2001).
→ Uma vez na célula bacteriana, o pH mais alto do citoplasma causa dissociação do ácido e a redução no pH da célula a qual interromperá reações enzimáticas e sistemas de transporte de nutrientes (Cherrington et al., 1991).
→ Isso se torna um grande problema para bactérias, uma vez que devem manter o pH do citoplasma próximo a neutralidade para sustentar suas funções macromoleculares.
→ Atuam provocando mudanças na permeabilidade e atividade da membrana celular das bactérias;
→ alterações na atividade dos canais de cálcio,
→ perturbação do equilíbrio iônico e
→ perda de íons.
A exportação de excesso de prótons requer consumo de adenosina celular trifosfatase (ATP), o que pode resultar na depleção de toda a energia celular levando a sua morte (Davidson, 2001), ou quando esta não ocorre, restará uma disponibilidade reduzida de energia para proliferação celular, o qual resultará em algum grau de bacteriostase (Mroz et al., 2000).
Atividades antibacterianas menos diretas (Figura 1) também foram atribuídas aos ácidos orgânicos e incluem interferência com transporte de nutrientes, danos na membrana citoplasmática em vazamentos, interrupção da permeabilidade da membrana externa, e influenciando a síntese macromolecular (Cherrington et al., 1991; Denyer e Stewart, 1998; Davidson, 2001).
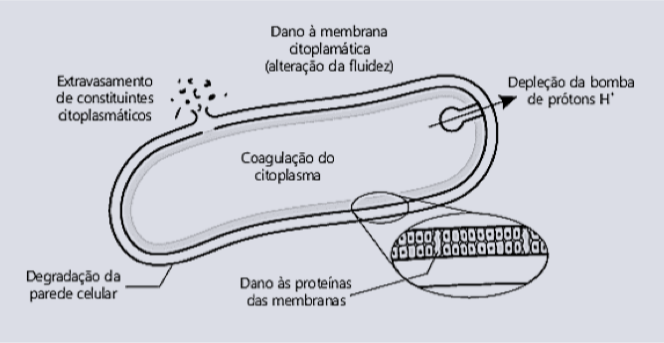

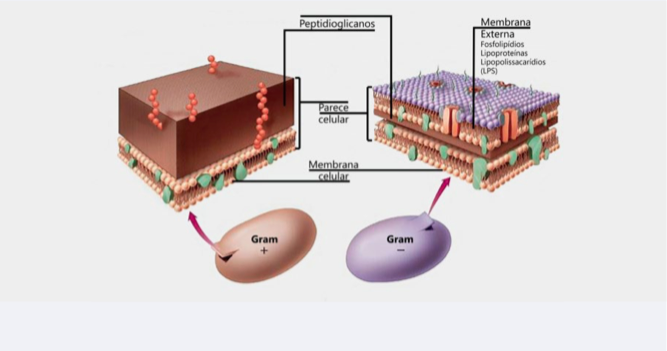
Os organismos Gram negativos possuem a membrana externa constituída de lipopolissacarídeos a qual envolve a parede celular e, conforme já comentado anteriormente, esta condição facilita a entrada dos ácidos graxos ao interior do citoplasma das células bacterianas.
Via de regra, a maioria das bactérias patogênicas de importância são Gram – (E.coli/ Salmonella) e as principais bactérias benéficas da microbiota intestinal são Gram + (Lactococcus, Lactobacillus; Bifidobacterium), as quais além de sobreviverem muito bem em meio ácido, elas mesmas também o produzem (lático/butírico).

O uso de antibióticos na produção animal está cada vez mais restrito, sendo que ácidos orgânicos são uma alternativa interessante.
→ aumento da secreção pancreática e efeitos tróficos na mucosa gastrointestinal,
→ melhor digestibilidade de proteínas e aminoácidos,
→ redução de amônia e produção de aminas biogênicas e
→ a modificação da microbiota intestinal.
Finalmente, há evidências de aumento/ crescimento da mucosa gastrointestinal na presença de ácidos orgânicos, particularmente ácidos graxos, como ácido butírico.
A falta de consistência na demonstração de um benefício do ácido orgânico está relacionada a variáveis não controladas, como capacidade de tamponamento dos ingredientes da dieta, presença de outros compostos antimicrobianos, limpeza do ambiente de produção e heterogeneidade da microbiota intestinal.

Archer, D. L. 1996. Preservation microbiology and safety: Evidence that stress enhances virulence and triggers adaptive mutations. Trends Food Sci. Technol. 7:91–95.
Bolduan, G., H. Jung, R. Schneider, J. Block, and B. Klenke 1988. Die wirkung von propion- und ameisensaure in der ferkelaufzucht J. Anim. Physiol A. Anim. Nutr. 59:72–78.
Burt, Sara. “Essential oils: their antibacterial properties and potential applications in foods—a review.” International journal of food microbiology 94.3 (2004): 223-253.
Cherrington, C. A., M. Hinton, and I. Chopra. 1990. Effect of short-chain organic acids on macromolecular synthesis in Escherichia coli. J. Bacteriol. 68:69–74.
Cherrington, C. A., M. Hinton, G. C. Mead, and I. Chopra. 1991. Organic acids: Chemistry, antibacterial activity and practical applications. Adv. Microb. Physiol. 32:87–108.
Cole, D. J. A., R. M. Beal, and J. R. Luscombe. 1968. The effect on performance and bacterial flora of lactic acid, propionic. Symposium acid, calcium propionate and calcium acrylate in the drinking water of weaned pigs. Vet. Rec. 83:459–464.
Davidson, P. M. 2001. Chap. 29. Chemical preservatives and natural antimicrobial compounds. Pages 593–627 in Food Microbiology—Fundamentals and Frontiers. 2nd ed. M. P. Doyle, L. R. Beuchat, and T. J. Montville, ed. American Society for Microbiology, Washington, DC.
Denyer, S. P., and G. S. A. B. Stewart. 1998. Mechanisms of action of disinfectants. Int. Biodeterior. Biodegradation 41:261–268.
Dibner, J. J. e Buttin, P. 2002. Use of Organic Acids as a Model to Study the Impact of Gut Microbiota on Nutrition and Metabolism. J. Appl. Poult. Res. 11:453–463
Eidelsburger, U., M. Kirchgessner, and F. X. Roth. 1992. Zum Einfluss von Fumarsaure, salzsaure, natriumformiat, Tylosin und Toyocerin auf tagliche zunahmen, futteraufnahme, futterverwertung und verdaulichkeit. J. Anim. Physiol. A. Anim. Nutr. 68:82–92.
Gedek, B., F. X. Roth, M. Kirchgessner, S. Wiehler, A. Bott, and U. Eidelsburger. 1992. Zum Einfluss von Fumarsaure, salzsaure, natriumformiat, Tylosin und Toyocerin auf die keimzahlen der microbiota und deren zusammensetzung in verschiedenen segmenten des gastrointestinaltraktes. J. Anim. Physiol. A. Anim. Nutr. 68:209–217
Jongbloed, A. W., Z. Mroz, R. van der Weij-Jongbloed, and P. A. Kemme. 2000. The effects of microbial phytase, organic acids and their interaction in diets for growing pigs. Livest. Prod. Sci. 67:113–122
Kemme, P. A., A. W. Jongbloed, Z. Mroz, J. Kogut, and A. C. Beynen. 1999. Digestibility of nutrients in growing-finishing pig is affected by Aspergillus niger phytase, phytate and lactic acid levels. 1. Apparent ileal digestibility of amino acids. Livest. Prod. Sci. 58:107–117.
Kemme, P. A., A. W. Jongbloed, Z. Mroz, J. Kogut, and A. C. Beynen. 1999. Digestibility of nutrients in growing-finishing pigs is affected by Aspergillus niger phytase, phytate and lactic acid levels. 2. Apparent total tract digestibility of phosphorus, calcium and magnesium, and ileal degradation of phytic acid. Livest. Prod. Sci. 58:119–127.
Matté, F. Melhoradores de desempenho naturais extratos herbais na produção animal. informativo técnico Vetanco – Brasil | nº 22 | fevereiro de 2019.
Mroz, Z. 2000. Supplementary organic acids and their interactive effects with microbial phytase in diets for pigs and poultry. Page 1 in Proc. Annu. Conf. Phytase in Anim. Nutr., Lublin, Poland.
Mroz, Z., A. W. Jongbloed, K. Partanen, K. Vreman, J. T. M. van Diepen, P. A. Kemme, and J. Kogut. 1997. The effect of dietary buffering capacity and organic acid supplementation (formic, fumaric, or n-butyric acid) on digestibility of nutrients (protein, amino acids, energy and minerals), water intake and excreta production in growing pigs. ID-DLO Rep. No. 97. 014. Institute for Animal Science and Health, Branch Runderweg, Lelystad, The Netherlands.
Nostro, A. et al. Susceptibility of methicilin-resistant Staphylococci to oregano essential oil, carvacrol, and thymol. FEMS Microbiology Letters, v. 230, n.3, p.191-195, 2004. Partanen, K. 2001. Organic acids—Their efficacy and modes of action in pigs. Page 201 in Gut Environment of Pigs. A. Piva, K. E. Bach Knudsen, and J. E. Lindberg, eds. Nottingham University Press, Nottingham, UK.
Ricke, S. C. 2003. Perspectives on the Use of Organic Acids and Short Chain Fatty Acids as Antimicrobials. Poultry Science 82:632–639.
Russell, J. B. 1992. Another explanation for the toxicity of fermentation acids at low pH: Anion accumulation versus uncoupling. J. Appl. Bacteriol. 73:363–370.
Scipioni, R., G. Zaghini, and B. Biavati. 1978. Ricerch sull’impiego di diete acidificate nello svezzamento precoce dei suinetti. Zool. Nutr. Anim. 4:201–218.

