

Neste segundo artigo vamos entender sobre os processos empregados para separação, concentração e os benefícios do uso aplicado dos constituintes das leveduras como ingrediente funcional na alimentação animal.
![]()
O seu fracionamento produz derivados que podem ser adicionados em dietas como a levedura autolisada, que abordamos no primeiro artigo (clique aqui se quiser ler o artigo na íntegra), além do extrato de levedura (citoplasma) e a parede celular (PC) que são obtidos pelo desmembramento da fração solúvel (extrato) e insolúvel (parede celular) das células autolisadas e que explicaremos com mais detalhes.


Após o processo de autólise, a biomassa é submetida a um fracionamento por centrifugação para obtenção da parede celular (Figura 1) e, em seguida, seca em spray dryer. Para garantir que o produto seja constituído apenas por parede celular, é necessário um controle continuado do processo para completa extração da parte solúvel.
Para isto, são realizadas quantas centrifugações forem necessárias, além da adição de ingredientes como cloreto de sódio que auxiliam na remoção e purificação da parede. Desta forma, as análises de proteína bruta (abaixo de 35%) durante o processo são procedimentos coadjuvantes para determinação e garantia de um produto constituído por 100% de parede celular, como é o caso do MAXIMOS.
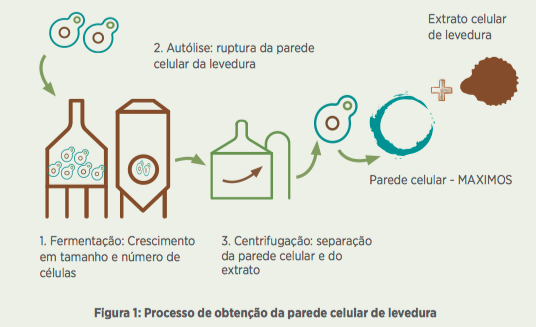

O papel principal da parede celular na levedura está relacionado à proteção física, ao controle osmótico e às propriedades de permeabilidade da célula (SMITH et al., 2000; KLIS et al., 2002).
![]()
![]()

De forma geral, a parede celular da levedura é composta por 85% de polissacarídeos e 15% de proteínas (BLASCOL et al, 2011).
A distribuição destes constituintes se organiza em duas camadas principais que são compostas basicamente por três macromoléculas: mananoproteína (20-23%), um complexo no qual o polissacarídeo manano está covalentemente ligado à proteína; glucana (48-60%), um polissacarídeo com ligações do tipo β-1,3 e β-1,6 de glicoses e quitina (0,6 – 2,7%), um polímero de ligações β-1,4 N-acetilglicosamina. (FLEET, 1985; HARTLAND et al., 1994; KLIS, 1994).

A camada externa, composta de mananoproteínas (MAP), representa de 20 a 23% do peso seco da parede celular. As cadeias de carboidratos, os mananoligossacarídeos, das MAP, são responsáveis pelo reconhecimento e interações célula-célula e com meio externo, além de serem capazes de determinar a especificidade imunológica de leveduras (COSTA et al., 2012).
As MAP são capazes de limitar a ação de enzimas exógenas como um mecanismo de defesa, conferindo resistência e proteção às células vivas de leveduras e limitando o acesso a todo seu conteúdo citoplasmático (COSTA et al., 2012), sendo estes um dos motivos pelos quais o processo de autólise (lise da parede celular) é tão importante para a obtenção de um produto com maior funcionalidade no animal.
A camada interna da parede celular possui uma rede tridimensional formada de cadeias β-(1,3)-glucana interligada a β-1,6-glucana e quitina, e confere a elasticidade e estrutura de parede celular de leveduras (KLIS et al., 2006).

Por sua vez, a β-(1,6)-glucana, corresponde à cerca de 5% do peso seco da parede celular e são moléculas relativamente pequenas, com cerca de 140 resíduos de glicose, atuando como apoio para a organização e integridade da camada de mananoproteínas (ROEMER et al., 1994).


As β-glucanas pertencem a uma classe conhecida como MRB (modificadores da resposta biológica), pois são capazes de alterar no animal a resposta biológica pela ação no sistema imune, auxiliando as células normais a produzir mensageiros químicos (mediadores) que melhoram a capacidade do sistema imunológico de identificar e produzir substâncias de defesa.
Esta atividade está relacionada ao tipo de ligações glicosídicas, peso molecular, conformação espacial, grau de polimerização e de ramificação (DIETRICHMUSZALSKA et al., 2011). Por exemplo, dentre as diferentes conformações, a tripla hélice é a que apresenta a maior atividade biológica (HA et al., 2002).
Assim, algumas moléculas de β-glucana podem ser capazes de modular a resposta imune desencadeando uma série de eventos como a produção de fatores quimiotáticos que induzem à migração de leucócitos para o sítio da infecção (LIU et al., 2008).
Também, o reconhecimento da β-glucana pelo sistema imune dos animais pode ocorrer por meio de receptores de superfície celular como dectin-I, já identificados em células imunes como macrófagos, monócitos e neutrófilos (BROWN e GORDON, 2005), além de estarem envolvidos na resposta imune específica pela expressão de diversas citocinas (LEE et al., 2001).

Além de serem classificadas como MRB, muitos trabalhos têm relacionado as β-glucanas à capacidade de se ligarem seletivamente às micotoxinas polares e apolares como zearalenona e aflatoxinas, através das ligações entre as pontes de hidrogênio e forças de Van der Waals.

Por meio de ensaios in vitro, Yiannikouris et al. (2003) relataram que este mecanismo ocorre devido à estrutura tridimensional da parede celular da levedura, que aumenta a superfície de contato das β-glucanas, de forma a expor um maior número de polos de ligação, aumentando a disponibilidade destes e, consequentemente, a eficiência em adsorver as micotoxinas.
Por consequência, a parede celular como o MAXIMOS apresenta uma função auxiliar de adsorção, sendo mais um mecanismo de apoio para a saúde animal.

O mananoligossacarídeo (MOS) presente na parede celular atua por meio de mecanismos de afinidade, sendo capaz de aderir-se às bactérias patogênicas no intestino do animal, impedindo que estas iniciem um processo de colonização, ou ainda modulando e preparando o sistema imunológico para proteção contra um processo infeccioso (SHANE, 2001).
Em relação ao primeiro mecanismo, sabe-se que para uma bactéria iniciar o processo infeccioso é necessário aderir-se aos receptores da superfície epitelial. As fímbrias manose-sensíveis (tipo 1) têm grande incidência entre as enterobactérias como Salmonella sp. e E. coli. tendo uma alta afinidade pela molécula do MOS.
Com isso, antes de se ligarem à mucosa intestinal, podem se aderir ao MOS presente no lúmen intestinal sendo assim eliminada junto ao bolo fecal (SHANE, 2001).

O MOS também pode aderir à mucosa intestinal, compondo uma barreira física de proteção que impede a colonização por microrganismos nocivos ao animal, proporcionando uma melhora na saúde e desempenho de animais.
Em adição a estes mecanismos, assim como ocorre com os β-glucanos, há evidências de que o MOS apresenta capacidade para induzir ativação de macrófagos por ocupar sítios receptores de manose nas glicoproteínas da superfície celular do macrófago (SILVA, 2006).
De acordo com Silva, et al., (2010), quando três ou mais destes sítios estão ocupados, inicia-se uma ativação da resposta imune adquirida que é caracterizada por uma reação em cascata que culmina na ativação dos macrófagos e na liberação de citocinas.
Assim, o MOS é capaz de aumentar os níveis de anticorpos circulantes específicos e a síntese de imunoglobulinas secretórias em resposta à exposição a antígenos (SAVAGE et al., 1997), sendo, na prática, efetivo no aumento de título vacinal, o que otimiza o efeito da vacinação e imunidade adquirida pelos animais imunizados.

Melhorias relacionadas à integridade intestinal dos frangos de corte e de aves de postura têm sido relatadas em estudos realizados pela Aleris utilizando o MAXIMOS (parede celular de S. cerevisiae).
![]()
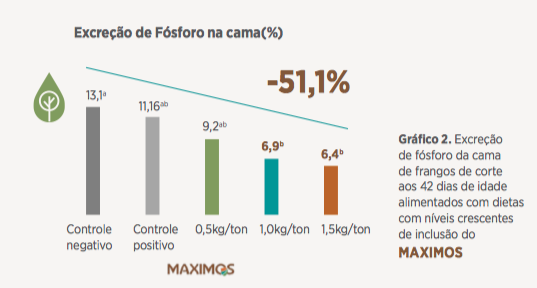
A capacidade do MOS em ampliar a área de absorção intestinal (ZAFAR et al., 2004), preservando as vilosidades pela diminuição da contaminação por patógenos e estímulo da microbiota benéfica, é um reflexo da saúde intestinal e melhor taxa de passagem verificadas neste experimento.



Neste mesmo experimento houve uma diminuição expressiva da excreção de fósforo na cama (Gráfico 2; – 51,1% para o nível de 1,5kg/t de Maximos) que segundo Fornazier et al., 2017 sinaliza uma melhora da saúde intestinal e melhor aproveitamento de nutrientes.
Com estes resultados, entende-se que o uso de parede celular como o MAXIMOS em dietas livres de antibióticos promotores de crescimento, que já é uma realidade no Brasil, assume uma função auxiliar no enfrentamento dos desafios entéricos e de ambiência comumente presentes nas criações.
A associação deste aditivo com outros de eficácia comprovada como acidificantes, fitoterápicos, probióticos, dentre outros, sustenta o status de saúde do animal e, consequentemente, seu desempenho. Entender as características e funcionalidade de cada aditivo é fundamental para assertividade e geração de bons resultados entendendo as especificidades de cada cliente final.

O extrato da levedura é a fração resultante da lise celular, após centrifugação e remoção da parede celular (fração insolúvel).
Muito rico nutricionalmente, o extrato integral da levedura, Saccharomyces sp., é descrito por Caballero-Córdoba et al., (1997) como uma excelente opção proteica não convencional para nutrição animal (Figura 2) devido ao teor elevado de proteína bruta (em média 48%, de acordo com AOAC), presença de ácidos nucleicos (em média, 8%, de acordo com método de HEBERT et al., 1971), lipídeos (em média, 4%, extraídos de acordo com o procedimento de BLIGHT e DYER, 1959), cinzas (em média, 8%, de acordo com AOAC) e carboidratos totais, que correspondem aos açúcares simples, oligossacarídeos, polissacarídeos, e seus derivados metabólicos (carboidratos de cadeias mais simples que podem ser resultados do metabolismo da levedura), incluindo éteres metílicos com grupos redutores livres ou potencialmente livres, (em média, 33%, método colorimétrico de DUBOIS et al.1956).

Com base neste conhecimento, durante muitos anos os produtos à base de levedura foram considerados como fontes de nucleotídeos para a nutrição animal.
Entretanto, apenas o processo de lise celular não é capaz de fornecer uma fonte padronizada de nucleotídeos livres.
Ou seja, só porque a levedura contém ácidos nucleicos não significa que estão biodisponíveis.
![]()
![]()
Assim, embora haja na literatura diversos estudos que relatem a utilização de nucleotídeos e atribuem as melhoras observadas nos animais à presença destes compostos, muitas vezes não são claras as informações sobre a fonte e disponibilidade dos nucleotídeos o que dificulta o entendimento sobre os reais benefícios e efeitos destas moléculas nos animais.

![]()
Os nucleotídeos são compostos intracelulares de baixo peso molecular que participam de vários processos bioquímicos e reações metabólicas, além de serem os constituintes (subunidades) dos ácidos nucleicos DNA e RNA (NELSON e COX, 2000).
Um nucleotídeo é formado por três moléculas que variam entre o DNA e o RNA, sendo:
![]()
![]()
![]()
O nucleosídeo é formado apenas pela base nitrogenada e a pentose, sem a ocorrência do grupo fosfato. A inosina é um exemplo de nucleosídeo que pertence ao grupo das purinas. Este nucleosídeo é formado pela ligação da hipoxantina a uma ribose (pentose).
Esta molécula é comumente encontrada em fitas de tRNA. O tRNA é originado a partir do processo de transcrição do DNA, sendo responsável por transportar os aminoácidos para a síntese proteica (NELSON e COX, 2000; RAVEN et al., 2014).
![]()
No entanto, hoje em dia, com os progressos em genética e ambiência para o máximo desempenho e, em determinadas fases em que o crescimento é acelerado ou o consumo de alimentos é insuficiente para atender as necessidades do animal, a suplementação de nucleotídeos é altamente recomendada para disponibilizar as bases nitrogenadas e nucleosídeos para serem utilizadas diretamente pelo organismo, economizando energia e/ou nutrientes adicionais que poderiam ser direcionados para outras funções.
Para que os nucleotídeos presentes nos ácidos nucleicos de ingredientes adicionados à dieta sejam absorvidos, é necessário que, primeiro, estes sejam hidrolisados em nucleosídeos no lúmen intestinal por ação de enzimas como as nucleases pancreáticas e fosfatases intestinais. (QUAN e UAUY, 1991).

Entretanto, por hidrólise parcial é possível retirar o radical fosfato dos nucleotídeos, não sendo este o fator determinante do aproveitamento pelo animal (NELSON e COX, 2000).
Também, pressupõe-se que há uma diferença no aproveitamento entre as bases purinas e pirimidinas pelo organismo, sendo que o intestino desempenha um papel importante na definição do destino metabólico das purinas da dieta e pouco é relatado sobre a regulação do metabolismo da pirimidina pelo enterócito.
Em uma revisão de Uauy et al., (1994) os autores relatam que a diferença entre purinas e pirimidinas está mais na forma como estas são metabolizadas do que de fato absorvidas.

Desta forma, a diferença na quantidade das bases presentes em um ingrediente parece não apresentar diferenças de aproveitamento pelo animal, uma vez que tanto os nucleosídeos como apenas as bases são absorvidas pelo intestino (QUAN e UAUY, 1991) e desempenham seu papel nas vias metabólicas.

De forma geral, no organismo animal, os nucleotídeos são sintetizados por duas vias metabólicas, a síntese de novo que ocorre em várias etapas, sendo um processo metabolicamente caro, em termos energéticos, por isso exige quantidades substanciais de energia na forma de ATP (CARVER et al., 1995) e a via de salvamento, que recicla as bases livres e os nucleosídeos liberados a partir da degradação de ácidos nucleicos (NELSON e COX, 2000).
Sob condições de alimentação regular, a via de salvamento recicla 90% ou mais das bases de purina.

![]()
Por exemplo, a mucosa intestinal, as células hematopoiéticas da medula óssea, leucócitos, eritrócitos e linfócitos são incapazes de realizar a via de síntese de novo (SANDERSON e HE, 1994) e, assim, utilizam a via de salvamento o que sugere que uma suplementação de nucleotídeos via dieta pode ser importante para essas células (COSGROVE, 1998).
Particularmente na mucosa intestinal, por ser um local de intenso turnover celular e, consequentemente, um alto gasto metabólico, a suplementação de nucleotídeos na dieta apresentam uma função significativa para a manutenção e otimização das atividades celulares.


Apesar dos nucleotídeos serem sintetizados endogenamente, a sua suplementação dietética pode exercer efeitos benéficos aos animais com ação no crescimento e desenvolvimento intestinal (WU et al., 2018) e estímulo a irrigação do tecido intestinal (BUSTAMANTE, 1990), além de acelerar a mitose e diminuir a apoptose dos enterócitos, devido à ausência (SAVAIANO e CLIFFORD, 1981) ou limitação (LELEIKO et al.,1983) da síntese de novo.
![]()

Um papel protetor também foi relatado por Martinez-Puig et al. (2007) na prevenção da diarreia em suínos, sendo que uma possível explicação é justamente a modulação da composição da microbiota fecal, por meio de um aumento da abundância dos gêneros favoráveis ao desempenho do animal (WU et al., 2018).
Outra função atribuída por Silva et al. (2013) aos nucleotídeos foi sua habilidade em promover taxas ótimas de divisão celular na epiderme, uma vez que garantem a rápida recuperação de qualquer dano a pele, e possuem capacidade de proteger as camadas subjacentes de danos, desidratação e patógenos, pois em um estudo utilizando ratos que receberam nucleotídeos de adenosina (principalmente ATP e ADP) houve evidências de que a presença destes nucleotídeos foi capaz de atenuar a resposta inflamatória dos animais com dermatite e acelerar a sua recuperação.
![]()

Baseado nos benefícios dos nucleotídeos, no conhecimento da riqueza do extrato de levedura e no empenho e esforço em se obter um produto que garanta nucleotídeos livres, a ALERIS, após anos de aprimoramento, apresenta em sua linha de produtos o NUCLEOBASE 1.5.
Este produto é originado do extrato de levedura que passa por um processo biotecnológico de purificação e hidrólise do RNA, incluindo, para a sua padronização, a levedura autolisada, com a garantia de uma quantidade mínima de 15% de nucleotídeos livres no produto final.


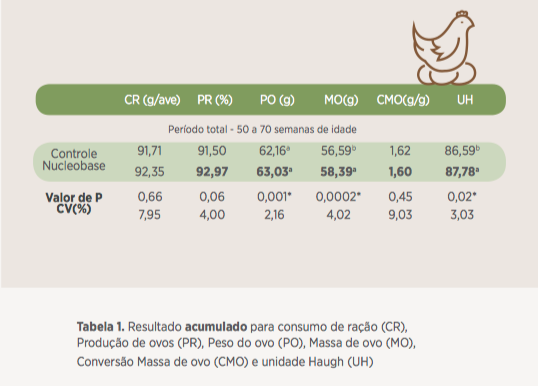
Ao final do período experimental as aves apresentaram melhora expressiva para taxa de postura (93% de postura, em média), peso do ovo (63 gramas, em média), deposição de massa de ovo (58 gramas/ave/dia, em média) e unidade Haugh (87,8 em média) quando alimentadas com dietas suplementadas com 500g/t de Nucleobase 1.5.
![]()


![]()








































