


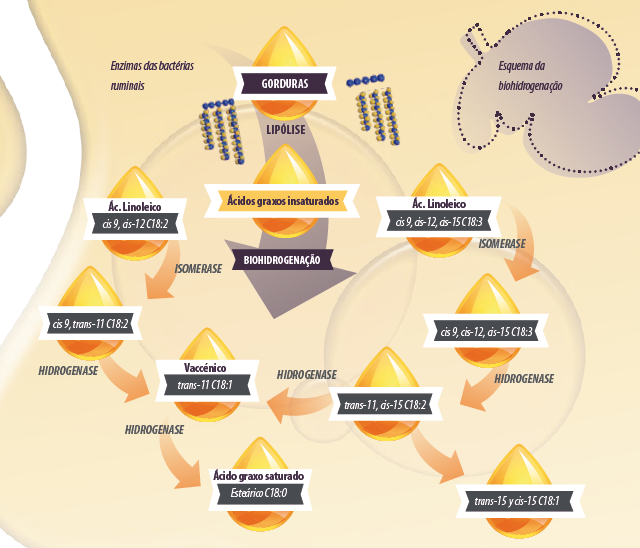
Nesta segunda parte do trabalho, analisaremos os resultados obtidos pelos pesquisadores nos ensaios in vivo e detalharemos com mais atenção a biohidrogenação das gorduras no rúmen, processo que, como já vimos, é a etapa posterior à lipolise, primeiro processo a que são submetidas as gorduras que entram no rúmen.
Este processo é considerado uma adaptação detoxificante (Kemp et al., 1984), que contribui marginalmente para a eliminação dos equivalentes redutores produzidos pela fermentação ruminal (Lourenço, et al. 2010). A biohidrogenação (BH) compreende várias etapas, dependendo dos AGI, e várias rotas, dependendo da dieta e ambiente ruminal (Griinari et al., 1998).

Além dos estudos baseados nos isolamentos selecionados, foram realizados esforços para avaliar in vivo a relação entre bactérias do rúmen e a biohidrogenação adicionando bactérias e medindo seus produtos. Ou, ainda, agregando suplementos dietéticos que, sabe-se, afetam a BH e medindo a abundância de bactérias. Em geral Os estudos in vivo de adição de gordura demonstraram que não há mudanças, ou diminuição nos gêneros de protozoários totais e principais.
⇑ Os estudos in vivo de adição de gordura demonstraram que não há mudanças, ou diminuição nos gêneros de protozoários totais e principais.
Outras observações:
- Hristov et el., 2012 observaram que o ácido láurico (C12:0) diminuiu fortemente a contagem de protozoários comparado aos ácidos mirístico (C14:0) e esteárico (C18:0).
- Oldick e Firkins (2000), de maneira mais geral, mostraram que ao aumentar o grau de insaturação diminui a contagem de protozoários, porém enfatizaram que esta mudança pode ser difícil de ser avaliada devido à variação individual, o que explica a inconsistência nos ensaios.
Observações sobre a comunidade arqueal
Os estudos com cepas puras de arqueas, adicionando ácidos orgânicos, ou ácidos graxos saturados, apresentaram inibição da produção de metano pelo Methanobrevibacter ruminantium.
- Lillis et al. (2011) comprovaram que a adição de óleo de soja invivo alterou a contagerm, porém, não a diversidade das arqueas metanogênicas. Eles levantaram a hipótese de que estas mudanças poderiam ser consequência da alteração do perfil de AGV (menos acetato e butirato, que produzem H2, além do propiônico), devido a mudanças na comunidade bacteriana.
- Hristov et al. (2012) sugeriram que mudanças na comunidade arqueal poderiam ser uma consequência da diminuição da abundância de protozoários com dietas ricas em gordura
Em cultivos puros, o ácido linoleico (AL) pode afetar o crescimento do fungo Neocallimastix frontalis (Maia et al., 2007).
Boots et al. (2012) confirmaram in vivo o efeito negativo da adição de AL na ordem Neocallimastix, cuja riqueza e diversidade diminuem com a adição de óleo de soja.
O efeito inibitório dos óleos sobre o crescimento bacteriano foi amplamente estudado em cultivos puros in vitro de cepas do rúmen (Maia et al., 2007), focando em bactérias que desempenham papel na fibrólise, amilólise e metabolismo das gorduras.
No geral, estes primeiros estudos sobre AG saturados e monoinsaturados enfatizaram que os efeitos das gorduras sobre as bactérias do rúmen dependem do metabolismo bacteriano, da insaturação dos AG e da configuração geométrica das ligações duplas.
Os efeitos negativos do AG sobre B. fibrisolvens são:
++ fortes para AG de cadeia longa eicosapentaenóico (EPA: cis-5, cis-8, cis-11, cis-14, cis017-C20:5) e os ácidos docosahexaenoico (DHA; cis-4, cis-7, cis-10, cis-13, cis-16, cis-19-C22:6).
De forma similar, ALA aumenta fortemente a fase de latência e diminui a taxa de crescimento de Propionibacterium acnesn (Maia et al., 2016).
Os efeitos dos suplementos de gordura foram pesquisados in vivo, estabelecendo bactérias por espécie a partir de PCR quantitativo (Martin et al., 2016; Vargas-Bello-Perez, et al., 2016) ou, por gênero, utilizando 16S rDNA pirosequenciamento (Zened et al., 2013a; Huws et al., 2014).
Estes experimentos basearam-se, principalmente, na adição de óleo, diferentemente da maioria dos estudos sobre cultivos de cepa pura, que utilizaram AG livres.
No geral, os efeitos foram menores que os observados com cultivos puros, o que poderia ser reputado ao tipo de gordura adicionada, ou ao fato de que os efeitos na fase de latência não podem ser vistos in vivo.
As mudanças na microbiota ruminal devido a uma maior proporção de concentrado são muito mais altas que os efeitos por adição de gorduras. Alguns gêneros também foram afetados de maneira diferente pela adição de óleo em dietas de baixo e alto concentrado, especialmente Acetitomaculum, lachnospira e Prevotella (Zened et al. 2011).
Entre os gêneros de bactérias, ou espécies estudadas em vários experimentos, a Fibrobacter e Ruminococcus foram afetados negativamente na maioria dos casos, porém os efeitos no Butyrivibio e Prevotella foram mais variáveis.
Hipótese nº2 Devendra e Lewis (1974) também propuseram que a gordura da dieta poderia diminuir a disponibilidade de íons para as bactérias devido à formação de sais, o que é consistente com os efeitos protetores das substâncias ricas em cálcio sobre a degradação da celulose quando se adiciona óleo à dieta (Brooks et al., 1954). No entanto, não pode explicar todos os efeitos negativos da gordura na dieta.
Hipótese nº3 Devendra e Lewis (1974) também atribuíram a hipótese de que AG poderia exercer toxicidade direta sobre as bactérias do rúmen, o que é consistente com a incorporação de AGI nos cultivos bacterianos (Bauchart et al. al. 1990). Esta toxicidade poderia ser consequência de um impedimento na passagem de nutrientes pela aderência dos AG à parede celular.
⇒ O ácido linoleico pode alterar a integridade celular, porém não existe uma relação entre esta alteração e o nível de inibição do crescimento bacteriano em diferentes cepas no rúmen, incluída B. fibrisolvens.
A relação entre os lipídeos da dieta e a microbiota do rumen está dominada pela toxicidade dos AGI em muitos microorganismos, especialmente as bactérias fibrolíticas.
- Sabe-se que o gênero Butyrivibrio está fortemente envolvido no processo de detoxificação
Muitos estudos recentes sugerem que as rotas bioquímicas são mais complexas e as bactérias envolvidas poderiam ser mais diversas do que se acreditava há várias décadas.
As aplicações práticas envolvem ambos os lados desta relação.
⇑ A adição de gordura à dieta dá forma à comunidade microbiana do rúmen, modula a função do rúmen e isto fez com que uma grande quantidade de pesquisas tenham se dedicado, nos últimos anos, à redução das emissões de metano, ainda que o modo de ação não seja compreendido por completo, podendo depender da fonte de gordura (Patra e Yu 2013).
⇓ Existem efeitos colaterais negativos na eficiência alimentar, como a redução do consumo de matéria seca (Beauchemin et al., 2009).
As opções mais adequadas para dar forma à microbiota do rúmen e sua atividade dependem de muitos fatores:
- Sistema de produção
- Condições econômicas
- Regulações locais, ou especificações
No entanto, para aplicar estas diferentes manipulações no campo, deve-se obter novos dados in vivo em diversas condições dietéticas, com estudos de longo prazo, porque a resiliência da microbiota ruminal, ou sua adaptação à degradação de compostos de plantas, pode alterar os efeitos no tempo (Weimer 2015).
Além disso, novas pesquisas aplicadas sobre o metabolismo ruminal das gorduras exige conhecer melhor quais microorganismos, mecanismos enzimáticos e interações entre os microorganismos e entre a microbiota e o hóspede estão envolvidos.
Autores: Fernando Bacha Baz (NACOOP) & Maria Jesús Villamide Díaz ETSIAAB _ Universidade Politécnica de Madri
Tenha acesso à primeira parte do artigo clicando aqui!