

Nos ruminantes, a maioria dos processos digestivos, especialmente a fermentação de carboidratos e degradação das proteínas da dieta, devem-se à microbiota ruminal: bactérias, protozoários, fungos e arqueas.

Esta microbiota simbiótica proporciona nutrientes ao hospedeiro, especialmente ácidos graxos voláteis (AGV), proteína microbiana e vitaminas, ainda que também ocorram perdas de energia devido à produção de metano.
Apesar da estabilidade ruminal por sua constância funcional e resiliência do ecossistema microbiano (Weimer et al, 2015), a microbiota ruminal apresenta grandes variações individuais (Jami y Mizrahi 2012) e pode ser alterada por mudanças dietéticas abruptas, ou importantes, por exemplo, de conteúdo no amido ou gordura.


A adição de gordura na dieta pode: Modular a função do rúmen, diminuindo as emissões de metano (Martin et al., 2016). Diminuir a ingestão e conteúdo de gordura no leite (Rabiee et al., 2012). Um limite para a suplementação das dietas de ruminantes com gorduras é seu efeito negativo sobre a degradabilidade ruminal, especialmente quando tem alto conteúdo de AGI (Brooks et al., 1954).
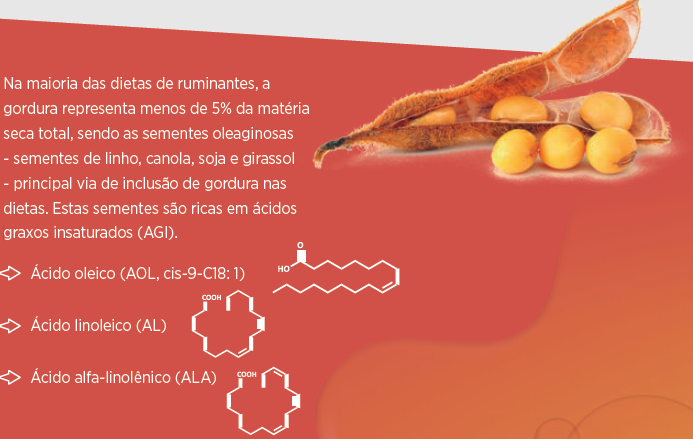
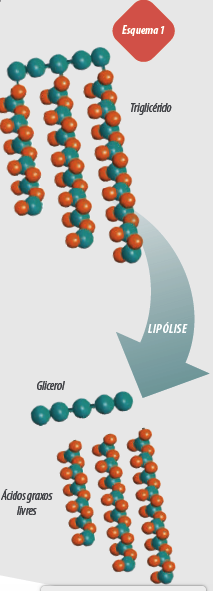
Neste trabalho vamos nos concentrar na influência da gordura sobre a microbiota ruminal. A maioria dos AG da dieta são ésteres de glicéridos: TRIACILGLICEROIS, principalmente, em concentrados GALACTOLIPÍDEOS e FOSFOLIPÍDEOS na forragem, exceto nas ensilagens nas quais as lipases vegetais liberam os ácidos graxos.


A biohidrogenação (BH) compreende vários passos, dependendo dos AGI, e várias rotas, dependendo da dieta e ambiente ruminal (Griinari et al., 1998).
Os fosfolipídeos e galactolipídeos podem ser hidrolizados por algumas cepas de Bacillus fibrisolvens (Hazlewood e Dawson 1979). Os tricilglicerois também são hidrolizados por diferentes espécies do grupo Butyrivibrio (Latham et al., 1972), porém a Anaereovibrio lipolyticus é a bactéria hidrolizadora de triglicéridos mais conhecida.
A lipase da Anaereovibrio lipolyticus foi estudada pela primeira vez por Henderson (1971) e seu genoma contém três genes que codificam lipases (Prove et al., 2013). As três enzimas foram mais ativas contra o láurico e o mirístico, que contra o palmítico, ou o esteárico, enquanto as gorduras dietéticas contém principalmente AG de 16 e 18 carbonos.









Estudos in vitro
Além dos estudos baseados nos isolamentos selecionados, foram realizados esforços para avaliar in vivo, ou in vitro a relação entre bactérias do rúmen e a biohidrogenação (BH) agregando bactérias e medindo seus produtos, ou agregando suplementos dietéticos que sabe-se afetam a BH e medir a abundância de bactérias.
Inoculando B. fibrisolvens no rúmen de cabras alimentadas com dieta enriquecida com óleo alto em ácido linoleico conjugado (CLA) total no fluido do rúmen, o que confirma que esta bactéria está envolvida na biohidrogenação in vivo (Shivani et al., 2016).

Pela maioria das gorduras dietéticas serem glicéridos e necessitarem sofrer uma lipólise antes da BH, a diminuição do metabolismo de AG no rúmen poderia limitar a BH no rúmen.
Um estudo preliminar com inibidores de esterase apresentou resultados promissores in vitro (Sargolzehi et al., 2015). Apás et al. (2015) mostraram uma proporção maior de cis-9, trans-11- CLA no leite de cabras suplementadas com uma mistura de Lactobacillus, Bifidobacterium y Enterococcus.
A gordura dietética dá forma à microbiota do rúmen, Brooks et al (1954) demonstraram que o óleo de milho, tanto in vitro como in vivo diminui a degradação da celulose ruminal e a produção de AGV, afetando a microbiota.
Também descobriram que a banha de porco, mais saturada que o óleo de milho, produz menor diminuição na degradação da celulose.
De maneira similar, Ikwuegbu y Sutton (1982) descobriram uma diminuição da degradabilidade da fibra, do percentual do acetato e do butirato, assim como um aumento do propinato quando usado o óleo de linhaça.
Os extratos de plantas também poderiam modular a atividade das bactérias biohidrogenadoras.
Os óleos essenciais diminuíram, ou aumentaram (Ishlak et al., 2015) a abundância de B. fibrisolvens in vitro, o que poderia explicar as mudanças no perfil dos produtos da BH no rúmen (Lourenço, et al. 2010). De maneira similar, os taninos diminuíram a abundância de B. proteoclasticus e aumentaram B. fibrisolvens (Ishlak et al., 2015) in vitro.






Por Fernando Bacha Baz (1) & María Jesús Villamide Díaz (2) (1)NACOOP, (2) ETSIAAB Universidade Politécnica de Madri
Acesse aqui a PARTE 2 deste artigo!