



Suplementação com arginina para fêmeas suínas em gestação: Parte I
A mortalidade pré-natal é um grande desafio ao desempenho reprodutivo de porcas hiperprolíficas, sendo este o maior problema reprodutivo dos mamíferos, incluindo humanos (Li et al., 2014).

(Kridli et al., 2016, Langendijk et al., 2016)

Além da mortalidade pré-natal, determinante no tamanho da leitegada, outros fatores como o peso ao nascimento do leitão e a uniformidade da leitegada são características de interesse econômico importantes.
Ademais, a hiperprolificidade tem sido associada ao crescimento intrauterino retardado (CIUR) em leitões. Quando a leitegada aumenta, o fluxo sanguíneo placentário também aumenta, contudo, em menor escala que o número de fetos, resultando em menor fluxo sanguíneo por feto e, portanto, menor suprimento de nutrientes e oxigênio (Père & Etienne, 2000).
A nutrição materna é o principal fator que afeta a sobrevivência, crescimento e desenvolvimento de embriões e fetos (Wu et al., 2006).
| Neste sentido, o uso de aminoácidos como nutriente funcional durante a gestação vem sendo largamente estudado devido à participação destes em vias metabólicas relacionadas às funções reprodutivas dos animais (Palencia et al., 2018). |
MECANISMOS DA ATUAÇÃO DA SUPLEMENTAÇÃO COM L- ARGININA NA RAÇÃO DE FÊMEAS SUÍNAS GESTANTES
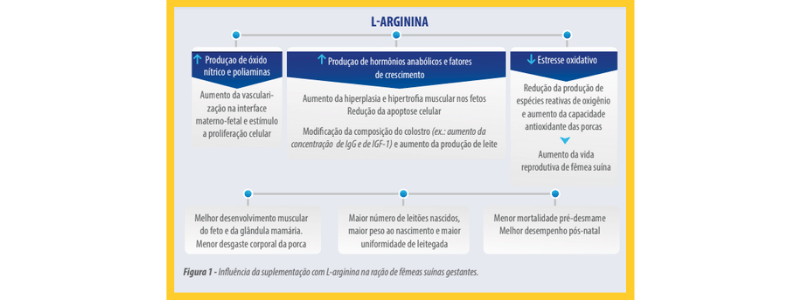
Dentre os aminoácidos utilizados como nutriente funcional, a arginina se destaca por, além da sua participação na síntese proteica no tecido muscular, ser precursor biológico de moléculas ativas como poliaminas (putrescina, espermina e espermidina) e óxido nítrico (NO) que favorecem a sobrevivência, o desenvolvimento e crescimento de embriões e fetos (Wu et al., 2013) através da promoção da proliferação celular e migração, angiogênese e dilatação de veias para aumento do fluxo sanguíneo (Wu et al.,2013; Krogh et al., 2017).
Considerando que o aporte de nutrientes para o útero ou para o feto é determinado tanto pela taxa de fluxo sanguíneo quanto pelas concentrações de nutrientes no sangue arterial e venoso (Wu et al, 2013; Wu et al., 2018), o desenvolvimento da vascularização do endométrio e da placenta são importantes na sobrevivência, crescimento e desenvolvimento do concepto (Bidarimath & Tayade, 2017).
As taxas de fluxo sanguíneo utero-placentária são altamente influenciadas pela disponibilidade de NO e de poliaminas (Reynolds et al. 2006; Khalil et al., 2015; Hussain et al., 2017). O NO e as poliaminas estimulam a proliferação e migração de células, remodelamento celular, angiogênese e a dilatação de vasos sanguíneos (Wu et al., 2006; Hsu &Tain, 2019)
A suplementação com arginina também pode reduzir a apoptose celular (Tan et al., 2010).
Estudos têm evidenciado melhora no desempenho reprodutivo de fêmeas suínas suplementadas com arginina principalmente no início da gestação (Wu et al., 2017), já que as maiores perdas embrionárias acontecem nesse período.
De fato, a formação da vascularização placentária se inicia logo após a implantação embrionária (Reynolds et al., 2006) com peso da placenta chegando ao platô, em média, aos 70 dias de gestação (Knight et al. 1977; Wu et al., 2005).[registrados]

| Porém o fluxo sanguíneo e a vascularização continuam a aumentar durante o terço final de gestação (Reynolds et al., 2006; Wu et al., 2013), assim sendo, a suplementação com arginina também pode ser benéfica nesse período (Quesnel et al., 2014). |
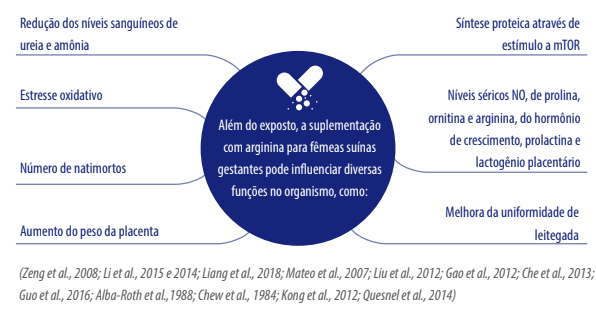
Considerando as variadas funções que a arginina e seus metabólitos (ornitina, poliaminas, prolina, glutamina, creatina e o NO) podem desempenhar na modulação do ambiente intrauterino (Figura 1), a suplementação com arginina para a fêmea suína gestante pode causar alterações epigenéticas com efeitos na anatomia, fisiologia e no metabolismo dos embriões e fetos (Kwon & Kim, 2017; Garcia et al., 2020).
As alterações epigenéticas podem afetar o desempenho do animal através da programação fetal. Os mecanismos relacionados a esses benefícios são complexos e interligados.
De forma breve, a programação fetal pode ser entendida como o resultado de mudanças específicas durante o desenvolvimento intrauterino que altera quantitativa e/ou qualitativamente a trajetória de desenvolvimento com resultados que perduram por toda vida do indivíduo.
Nesse contexto, o abrangente estudo da influência dos nutrientes em todo o genoma é chamado nutrigenômica.
Em estudo de Zeng et al. (2008), a suplementação com 1,3% de L-arginina HCl na ração de ratos resultou em aumento do número de fetos e no aumento da expressão da enzima óxido nítrico sintetase induzível (iNOS) e endotelial (eNOS) nos sítios de implantação, evidenciando o efeito benéfico do NO na sobrevivência embrionária.
De fato, a suplementação com arginina no terço inicial de gestação promoveu aumento da produção sistêmica de NO, indicada pela elevação de seus produtos finais estáveis (nitrito e nitrato) no plasma de matrizes suínas gestantes quando suplementadas com 1,3% de arginina (Li et al., 2015), do número de vasos e do peso da placenta em 21 e 34% quando suplementadas com, respectivamente, 0,4 e 0,8% de arginina (Li et al., 2014).
Os principais genes que modulam os processos de vasculogênese e angiogênese na placenta e que podem ser afetados pela suplementação com arginina incluem:
- Fator de crescimento placentário 1 (PGF1);
- Famílias de proteínas do fator de crescimento endotelial (VEGF);
- Angiopoietinas (ANGT1, ANGT2);
- Fator de crescimento fibroblástico 2 (FGF2), assim como os seus respectivos receptores (Reynolds et al., 2010).
A expressão do VEGF e o seu receptor (VEGFR2), o qual é regulado positivamente em resposta à expressão de VEGF, é vital para adequada angiogênese (Otrock et al., 2007).
O VEGF é considerado o mais potente estimulador da angiogênese, promovendo o aumento no número de vasos sanguíneos na interface placentaendométrio em suínos (Vonnahme & Ford., 2004) e em tecidos em desenvolvimento (Otrock et al., 2007).
O VEGF é um potente fator de sobrevivência endotelial também conhecido por induzir a vasodilatação pelo aumento de NO (Liu et al., 2012). Além das poliaminas e do NO, a arginina é precursora de ornitina, ureia e creatina. Através do catabolismo da creatina, a arginina pode atuar no metabolismo da metionina e afetar a metilação do DNA (Robles et al., 2019).
A síntese de creatina a partir do guanidoacetato (produzido pela ação da enzima glicina-aminotransferase na arginina) ocorre através da transferência de um grupo metil do doador S-adenosilmetionina (SAM). A SAM também está envolvida no metabolismo da metionina como um doador de metil e seu uso para a síntese de creatina pode diminuir o pool de metil para a metilação do DNA (Robles et al., 2019).
Tem sido demonstrado que a expressão de alguns microRNAs podem afetar a angiogênese e vascularização (Hua et al. 2006; Poliseno et al., 2006; Anand et al., 2010). Causando o bloqueio da migração da célula endotelial, proliferação e angiogênese e ainda, regulando indiretamente a expressão da enzima e NOS (Poliseno et al. 2006).
Liu et al. (2012) demonstraram que a suplementação com arginina afeta a expressão de microRNAs envolvidos nesses processos (miRNA-15b e miRNA-222), podendo controlar a expressão do VEGFA e da eNOS.
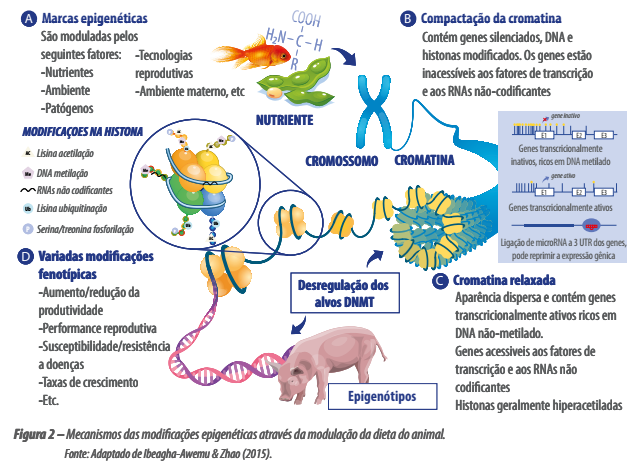
A Figura 2 demonstra como ocorrem as alterações epigenéticas relacionadas a nutrientes ingeridos na dieta.
Em decorrência disso, o entendimento dos processos relacionados ao desenvolvimento fetal em animais de produção tem sido alvo de pesquisas recentes (Kong et al., 2012; Zhou et al, 2016; Muns et al., 2016; Zhu et al., 2018; Chen et al., 2018; Costa et al., 2019; Garcia et al., 2020).
A partir desse conhecimento, torna-se possível a adoção de estratégias alimentares durante os diferentes estágios da gestação de fêmeas suínas com objetivos específicos.
CONSIDERAÇÕES FINAIS
Além do papel regulador na composição epigenética do indivíduo alimentado, os nutrientes, em determinada quantidade e qualidade, também podem afetar o desempenho da progênie.
Neste sentido, estratégias nutricionais, como a suplementação com arginina durante a gestação de fêmeas suínas, podem ser fator chave na melhoria da produtividade, principalmente em sistemas de produção com ciclo reprodutivo rápido e alto número de descendentes, como o de suínos (Murdoch et al., 2016).
O estudo da nutrigenômica pode ser implementado na seleção de nutrientes e identificação de marcadores relacionados a modulação da expressão gênica e do DNA, objetivando melhorias em características desejáveis.
Como por exemplo, na saúde, reprodução e produtividade do animal, tanto em condições ambientais adequadas quanto em situações de desafio.
Embora vários estudos demonstrem que a suplementação com arginina na ração de fêmeas suínas gestantes melhora o seu desempenho reprodutivo, ainda permanece em grande parte elusivo os mecanismos subjacentes pelos quais esse fato acontece. Ademais, não há definição clara ou consenso de qual o melhor período ou períodos e doses de suplementação com esse aminoácido durante a gestação
Referências

Bidarimath, M.; Tayade, C. Pregnancy and spontaneous fetal loss: A pig perspective. Molecular Reproduction and Development, v. 84, p. 856–69. 2017. doi: https://doi.org/10.1002/mrd.22847.
Che, L.; Yang, P.; Fang, Z.; et al. Effects of dietary arginine supplementation on reproductive performance and immunity of sows. Czech Journal of Animal Science, v. 58, p. 167–175. 2013. doi: https://doi.org/10.17221/6711-cjas.
Chen, X.; Guo, Y.; Jia, G.; et al. Arginine promotes slow myosin heavy chain expression via Akirin2 and AMPK signaling pathway in porcine skeletal muscle satellite cells. Journal of Agricultural and Food Chemistry, v. 18, p. 4734-4740. 2018. doi: https://doi.org/10.1021/acs.jafc.8b00775
Chew, B. P.; Eisenman, J. R.; Tanaka, T.S. Arginine infusion stimulates prolactin, growth hormone, insulin, and subsequent lactation in pregnant dairy cows. Journal of Dairy Science, v. 67, p. 2507–2518. 1984. doi: https://doi.org/10.3168/jds.S0022-0302(84)81607-0.
Gao, K.; Jiang, Z.; Lin, Y.; et al. Dietary l-arginine supplementation enhances placental growth and reproductive performance in sows. Amino Acids, v. 42, p. 2207–2214. 2012. doi: https://doi.org/10.1007/s00726-011-0960-9.
Garcia, I.S.; Teixeira, S.A.; Costa, K.A.; et al. L-Arginine supplementation of gilts during early gestation modulates energy sensitive pathways in pig conceptuses. Molecular Reproduction and Development, v. 87, p. 819–834. 2020. doi: https://doi.org/10.1002/mrd.23397
Guo PA, Jiang ZYA, Gao KGB, Wang LB, Yang XFB, Hu YJB, et al. Diets in Pregnancy Increases the Total and Live-Born Litter Sizes in Gilts. Anim Prod Sci 2012;57(6) 1091. doi: https://doi.org/http://dx.doi.org/10.1071/AN15156.
Hsu CN, Tain YL. Impact of arginine nutrition and metabolism during pregnancy on offspring outcomes. Nutrients 2019;11:1–15. doi: https://doi.org/10.3390/nu11071452.
Hua, Z.; Lv, Q.; Ye, W.; et al. MiRNA-directed regulation of VEGF and other angiogenic factors under hypoxia. PLoS One, v.;1(1), e116. 2006. doi:10.1371/journal.pone.0000116.
Hussain, T.; Tan, B.; Ren, W.; et al. Exploring polyamines: Functions in embryo/fetal development. Animal Nutrition, v. 3, p. 7–10. 2017. doi: https://doi.org/10.1016/j.aninu.2016.12.002.
Ibeagha-Awemu, E. M.; Zhao X. Epigenetic marks: regulators of livestock phenotypes and conceivable sources of missing variation in livestock improvement programs. Frontiers in Genetics, v. 6, artigo 302. 2015. doi: https://doi.org/10.3389/fgene.2015.00302
Khalil, A.; Hardman, L.; O’Brien, P. The role of arginine, homoarginine and nitric oxide in pregnancy. Amino Acids, v. 47, p. 1715–1727. 2015. doi: https://doi.org/10.1007/s00726-015-2014- 1.
Knight, J. W.; Bazer, F. W.; Thatcher, W. W.; et al. Conceptus development in intact and unilaterally hysterectomized-ovariectomized gilts: interrelations among hormonal status, placental development, fetal fluids and fetal growth. Journal of Animal Science, v. 44, p. 620–637. 1977. doi: https://doi.org/10.2527/jas1977.444620x.
Kong, X.; Tan, B.; Yin, Y.; et al. l-Arginine stimulates the mTOR signaling pathway and protein synthesis in porcine trophectoderm cells. Journal of Nutritional Biochemistry, v. 23, p. 1178-1183. 2012. doi: https://doi.org/10.1016/j.jnutbio.2011.06.012.
Kridli, R. T.; Khalaj, K.; Bidarimath, M.; et al. Placentation, maternal-fetal interface, and conceptus loss in swine. Theriogenology, v. 85(1), p. 135-44. 2016. doi: 10.1016/j.theriogenology.2015.08.001.
Krogh, U.; Oksbjerg, N.; Storm, A. C.; et al. Mammary nutrient uptake in multiparous sows fed supplementary arginine during gestation and lactation. Journal of Animal Science, v.95, p. 2517–2532. 2017. doi: https://doi.org/10.2527/jas2016.1291.
Kwon, E. J.; Kim, Y. J. What is fetal programming?: a lifetime health is under the control of in utero health. Obstetrics & Gynecology Science, v. 60(6), p. 506-519. 2017. doi: 10.5468/ogs.2017.60.6.506.
Langendijk, P.; Chen, T. Y.; Athorn, R. Z.; et al. Embryonic survival at day 9, 21 and 35 of pregnancy in intact and unilaterally oviduct ligated multiparous sows. Animal, v. 8, p. 1336-41. 2016. doi: 10.1017/S175173111600029X.
Li, J.; Xia, H.; Yao, W.; et al. Effects of arginine supplementation during early gestation (day 1 to 30) on litter size and plasma metabolites in gilts and sows, J. Anim. Sci. v.93, p. 5291–5303. 2015. doi: https://doi.org/10.2527/jas.2014-8657.
Li, X. L.; Bazer, F. W.; Johnson, G. A.; et al. Dietary supplementation with l-arginine between days 14 and 25 of gestation enhances embryonic development and survival in gilts. Amino Acids, v. 46, p. 375–384. 2014. doi: https://doi.org/10.1007/s00726-013-1626-6.
Liang, M.; Wang, Z.; Li, H.; et al. L-Arginine induces antioxidant response to prevent oxidative stress via stimulation of glutathione synthesis and activation of Nrf2 pathway. Food and Chemical Toxicology, v. 115, p. 315–328. 2018. doi: https://doi.org/10.1016/j.fct.2018.03.029.
Liu, X.D.; Wu, X.; Yin, Y.L.; et al. Effects of dietary l-arginine or N-carbamylglutamate supplementation during late gestation of sows on the miR-15b/16, miR-221/222, VEGFA and eNOS expression in umbilical vein Amino Acids, v.42, p.2111. 2012. doi: https://doi.org/10.1007/s00726-011-0948-5.
Mateo, R. D.; Wu, G.; Bazer, F. W.; et al. Dietary l- arginine supplementation enhances the reproductive performance of gilts. Journal of Nutrition, v.137, p. 652–656. 2007. doi: https://doi.org/10.1093/jn/137.3.652.
Muns, R.; Manzanilla, E. G.; Sol, C.; et al. Piglet behavior as a measure of vitality and its influence on piglet survival and growth during lactation. Journal of Animal Science, v. 91(4), p. 1838-43. 2013. doi: 10.2527/jas.2012-5501.
Murdoch, B. M.; Murdoch, G. K.; Greenwood, S.; et al. Nutritional influence on epigenetic marks and effect on livestock production. Frontiers in Genetics, v. 7, p. :1–10. 2016. doi: https://doi.org/10.3389/fgene.2016.00182.
Otrock, Z. K.; Makarem, J.A.; Shamseddine, A.I. Vascular endothelial growth factor family of ligands and receptors. Blood Cells, Molecules and Diseases, v. 38, p. 258–68. 2007. doi: https://doi.org/10.1016/j.bcmd.2006.12.003
Palencia, J. Y. P.; Lemes, M. A. G.; Garbossa, C. A. P.; et al. Arginine for gestating sows and foetal development: A systematic review. Journal of Animal Physiology and Animal Nutrition, v. 102(1), p. 204-213. 2018. doi: 10.1111/jpn.12679.
Père, M. C.; Etienn, M. Uterine blood flow in sows: Effects of pregnancy stage and litter size. Reproduction Nutrition Development, v. 40, p. 369–382. 2000. doi: 10.1051/rnd:2000105
Poliseno, L.; Tuccoli, A.; Mariani, L.; et al. MicroRNAs modulate the angiogenic properties of HUVECs. Blood. 2006 Nov 1;108(9):3068-71. doi: 10.1182/blood-2006-01-012369.
Quesnel, H. N.; Roy, Q.H.; Lottin, A.; et al. Supplying dextrose before insemination and L-arginine during the last third of pregnancy in sow diets: Effects on within-litter variation of piglet birth weight. Journal of Animal Science, v.92, p. 1445–1450. 2014. doi: https://doi.org/jas.2013-6701 [pii];10.2527/jas.2013-6701.
Reynolds, L. P.; Borowicz, P. P.; et al. Uteroplacental vascular development and placental function: an update. The International Journal of Developmental Biology, v. 54(2-3), p. 355-366. 2010. doi: 10.1387/ijdb.082799lr.
Reynolds, L. P.; Caton, J. S.; Redmer, D. A.; et al. Evidence for altered placental blood flow and vascularity in compromised pregnancies. The Journal of Physiology, v. 572, p. 51–58. 2006. doi: https://doi.org/10.1113/jphysiol.2005.104430.
Tan, B.; Yin, Y.; Kong, X.; et al. L-Arginine stimulates proliferation and prevents endotoxin-induced death of intestinal cells. Amino Acids, v. 38, p. 1227–35. 2010. doi: https://doi.org/10.1007/s00726-009-0334-8.
Vonnahme, K. A.; Ford, S. P. Placental vascular endothelial growth factor receptor system mRNA expression in pigs selected for placental efficiency. Jornal of Physiology, v. 1;554(Pt 1), p. 194-201. 2004. doi: 10.1113/jphysiol.2003.055061.
Wu G, Bazer FW, Johnson GA, Herring C, Seo H, Dai Z, et al. Functional amino acids in the development of the pig placenta. Molecular Reproduction and Development, v. 84, p. 870–882. 2017. doi: https://doi.org/10.1002/mrd.22809
Wu, G.; Bazer, F. W.; Johnson, G. A.; et al. Board-invited review: Arginine nutrition and metabolism in growing, gestating, and lactating swine. Journal of Animal Science, v. 96, p. 5035–51. 2018. doi: https://doi.org/10.1093/jas/sky377.
Wu, G.; Bazer, F. W.; Wallace, J. M.; et al. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. Journal of Animal Science, v. 84, p. 2316–37. 2006. https://doi.org/10.2527/jas.2006-156.
Wu, G.; Bazer, F.W.; Satterfield, M.C.; et al. Impacts of arginine nutrition on embryonic and fetal development in mammals. Amino Acids. v.45, p.241–256. 2013. doi: https://doi.org/10.1007/s00726-013-1515-z.
Zeng, X.; Wang, F.; Fan, X.; et al. Dietary arginine supplementation during early pregnancy enhances embryonic survival in rats. Journal of Nutrition, v. 138, p.1421–1425. 2008. doi: https://doi.org/10.1093/jn/138.8.1421.
Zhu, Y.; Li, T.; Huang, S.; et al. Maternal L-glutamine supplementation during late gestation alleviates intrauterine growth restriction-induced intestinal dysfunction in piglets. Amino Acids, v. 50(9), p. 1289-1299. 2018 doi: 10.1007/s00726-018-2608-5.
Zou, T.; Yu, B.; Yu, J.; et al. Moderately decreased maternal dietary energy intake during pregnancy reduces fetal skeletal muscle mitochondrial biogenesis in the pigs. Genes & Nutrition, v. 23, p. 11:19. 2016. doi: 10.1186/s12263-016-0535-1. PMID: 27551320; PMCID: PMC4968452. [/registrados]