



Utilização de glicose em ruminantes
Diferenças entre a absorção e o metabolismo do amido e da glicose em ruminantes
| METABOLISMO DE GLICOSE EM RUMINANTES |

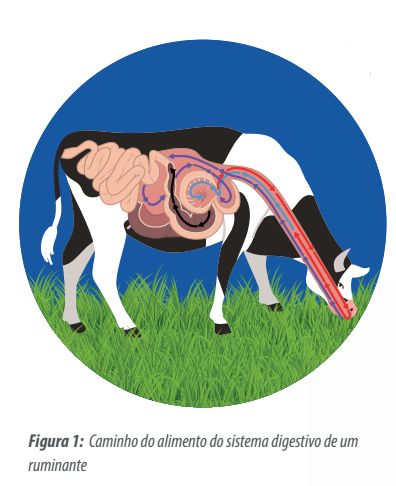
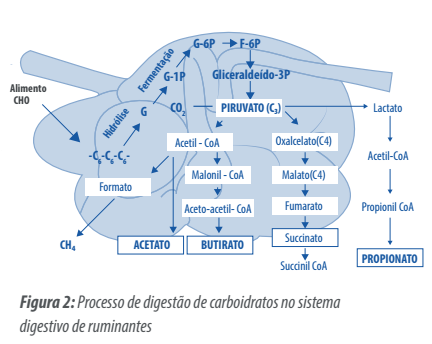
Embora o intestino delgado dos ruminantes esteja preparado para absorver glicose, geralmente, esse carboidrato não atinge o intestino porque fermenta rapidamente no rúmen (NRC. 2001).

Nesse processo de fermentação ruminal, a glicose é transformada no ácido graxo volátil de 3 carbonos – ácido propiônico – que é absorvido pelas paredes do rúmen e transformado em glicose no fígado (Mc Donalds, 1995)

| METABOLISMO DO AMIDO EM RUMINANTES |
O caso do amido é diferente. Seu metabolismo depende de:
![]()
![]()
![]()
Owens e Zinn calculam que sua digestibilidade no trato digestivo total em vacas leiteiras e confinadas é entre 92 e 98% (Owens e Zinn, 2005).


Para calcular a energia fornecida pelo amido como energia líquida, é necessário saber onde ele é digerido, onde é degradado e absorvido pelo animal, no rúmen ou no intestino delgado (Harmon e McLeod 2001).
A contribuição da fermentação do amido através do intestino grosso é menor, pois o amido que passou por todo o animal até atingir o cólon é, geralmente, menos degradável.
| DIGESTÃO E ABSORÇÃO DA GLICOSE |


No entanto, as limitações na digestão do amido e a absorção de glicose pelo intestino delgado muitas vezes desperdiçam essa maior eficiência (Mac Leod, 2001).
Huntington (2006), em uma tentativa de prever a digestibilidade do amido que entra no intestino delgado, traçou uma curva variando de 88% a 44% de digestibilidade quando a quantidade de amido que entra no intestino delgado oscila entre 300 a 1,5kg (Huntington 2006) .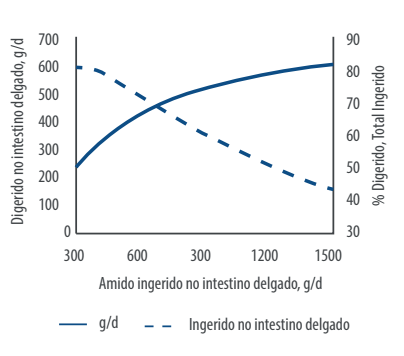
Mc Leod em 2007, comparando o amido com a glicose como fontes de energia, em um de seus experimentos, infundiu 775 gr / dia de amido hidrolisado e 900 gr / dia de glicose, tanto no intestino como no rúmen, em animais com 280 kg de peso vivo mantendo uma dieta basal.



A razão pela qual ocorre essa diferença entre as fontes de energia não é conhecida, uma vez que os níveis de insulina nos animais não variaram entre os tratamentos, mas duas hipóteses são especuladas:
- O excesso de glicose que atua como um ativador da lipogênese nas células.
- Efeito causado pelo excesso de acetato que atua na síntese de novo dos ácidos graxos, estimulando-a.
[registrados]Ao contrário da glicose, a absorção de ácidos graxos voláteis ocorre através das paredes do rúmen sem um gasto de energia para o animal. Já a absorção de glicose requer um gasto de energia para ativar o sistema de transporte (GLUT 4) através da parede intestinal.
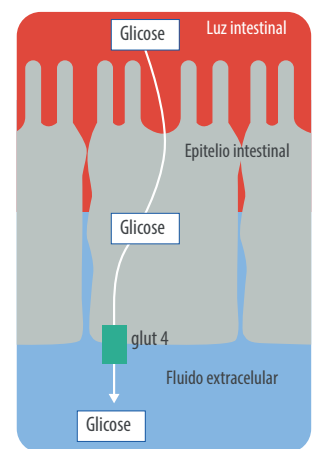

Ocorre também um gasto de enzimas, como a maltase, para quebrar a maltose em glicose e então absorvê-la (Baldwin 1968, 1995). Por mais que se gaste energia na absorção de glicose através do intestino delgado, ela não compensa a diferença dos 4 ATP para converter o propionato em glicose no fígado em relação à absorção de glicose diretamente do intestino delgado (Mc Donalds 1995).
Quando a glicose é absorvida, os tecidos esplênicos (baço) e, principalmente, toda a víscera portal têm um efeito duplo;
- Por um lado não só absorvem como consomem grande parte do que é absorvido e,
- Por outro lado, secretam hormônios como a insulina ou o glucagon.
A título de exemplo, a víscera portal é grande consumidora de glicose, ácidos graxos voláteis e aminoácidos (Blouin, 2002).
| RELAÇÃO ENERGIA E RETENÇÃO NITROGÊNIOGLICOSE NA CARNE E NO LEITE |
A quantidade de glicose absorvida no intestino que não é consumida pelos tecidos esplênicos e passa para a corrente sanguínea para ser utilizada pelos músculos ou glândula mamária dependerá do tipo e do estado fisiológico do animal testado, e das condições ambientais ao qual esse animal está sujeito.


Estudos sugerem que a vaca apresenta uma grave deficiência de glicose logo após o parto. Isto somado ao menor consumo de matéria seca e a alta demanda de nutrientes da lactação, desencadeia diferentes mecanismos metabólicos desenvolvidos pela vaca para lidar com a falta de glicose.
A ativação de todos os mecanismos metabólicos para fabricar glicose de vários precursores, tais como aminoácidos, glicerol, lactato e propionato é um deles.
- Aminoácidos
- Glicerol
- Lactato
- Propionato
A resistência à insulina é outra, mas essa resistência é seletiva, já que o feto e a glândula mamária não dependem da ação da insulina para a captação de glicose, tornando-os tecidos receptores da maior parte da glicose que é produzida ou entra no corpo (Reynolds, 2001).
Está bem demonstrado que elevar a concentração de glicose no sangue após o parto é fundamental para conseguir uma boa reprodução. Em seu trabalho, Garverick et al., 2013 obtiveram melhor taxa de prenhez no primeiro serviço com vacas que apresentavam maiores concentrações de glicose no sangue.

Os autores sugerem que apenas 50% da perda de produção devido ao estresse térmico seria explicada pela diminuição no consumo, e que os 50% restantes seriam explicados por alterações metabólicas na vaca.
Aparentemente, a vaca que sofre por estresse calórico é impedida de mobilizar ácidos graxos não esterificados do tecido adiposo, não podendo utilizá-los como fonte de energia para compensar a falta de
consumo.
A possível explicação para a falta de mobilização dos tecidos gordurosos seriam os altos níveis de insulina encontrados no sangue, visto que a insulina é um hormônio antilipolítico potente (O’Brien, 2010). Isso, somado à dessensibilização à insulina de quase todos os tecidos, exceto a glândula mamária e o feto, representaria uma oportunidade para que a glicose que o animal fabrica ou ingere possa ser usada diretamente para produzir leite e
emprenhar.
Russi et al (2011) demonstraram claramente em seu ensaio com vacas de transição que o aumento dos níveis de glicose no sangue produziu mais lactose no leite e melhor condição corporal para vacas que consumiram suplemento de glicose protegido da degradação ruminal.
Usando o mesmo suplemento, Mcarthy et al (2020) detectaram menor concentração de ácidos graxos não esterificados no sangue, beta hidroxibutirato, e marcadores inflamatórios, como LPB e haptoglobina. Russi et al (2011) demonstraram claramente em seu ensaio com vacas de transição que o aumento dos níveis de glicose no sangue produziu mais lactose no leite e melhor condição corporal para vacas que consumiram suplemento de glicose protegido da degradação ruminal.
Usando o mesmo suplemento, Mcarthy et al (2020) detectaram menor concentração de ácidos graxos não esterificados no sangue, beta hidroxibutirato, e marcadores inflamatórios, como LPB e haptoglobina.

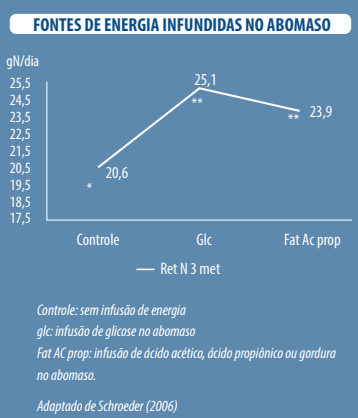
Em um de seus experimentos, Schroeder (2006) usou diferentes fontes de energia infundidas no abomaso ou no rúmen para avaliar a eficiência de utilização do N:
- Abomaso, glicose a uma taxa de 360 g/dia
- Rúmen: ácidos acético, propiônico e butírico
Todos os excessos de energia tornaram o uso do nitrogênio mais eficiente sem diferenças significativas, mas numericamente a glicose foi a fonte de energia que produziu com maior eficiência.
Eficiência no uso do nitrogênio com diferentes fontes de energia infundidas no abomaso
Russi, el al (2014) utilizou glicose protegida na criação de bezerros e observou que aqueles que consumiram 180 g de glicose protegida ao longo do ciclo de engorda tiveram menor consumo e melhoraram a eficiência de conversão.

Apesar do menor consumo por animal, não foram observadas diferenças nos parâmetros de concentração de glicose no sangue ou de insulina, ácidos graxos não esterificados ou ureia plasmática, sugerindo que os animais que consumiram 180 g de glicose protegida apresentaram metabolismo mais eficiente.
Em seu experimento, Mac Leod (2007) infundiu amido hidrolisado ou glicose, tanto no intestino como no rúmen, e uma das conclusões desse estudo foi que a alimentação com glicose infundida no intestino aumentou o acúmulo de tecido adiposo em todo o animal, especialmente, no tecido omental.
Vantagens também podem ser encontradas quando os animais na fase de terminação são alimentados com glicose protegida durante o verão, já que, aparentemente, o mesmo mecanismo de menor mobilização do tecido adiposo que atua em vaca em lactação estaria agindo em bovinos de corte (O’Brien, 2010).

Em seu experimento, Russi (2019) utilizou glicose protegida como suplemento para alimentar novilhos em terminação durante o verão onde o Índice de Temperatura e Umidade (ITU) foi de 72 ± 4,9. A resposta ao suplemento tornou-se mais evidente nos picos de estresse térmico, em que animais que se alimentaram com glicose consumiram menos, mas tiveram mesmo peso vivo, mostrando uma melhor eficiência de conversão.
Em vacas reprodutoras, Petersen et al descobriram que alimentar com precursores gliconeogênicos, como proteína bypass ou como sais de propionato, permite reduzir os dias até o primeiro estro e % de prenhez, mudando a condição corporal e o peso do bezerro ao nascimento em vacas de 2 ou 3 anos.
| CONCLUSÃO |

