





Un estudio reciente sobre las tasas de mortalidad en Alemania de terneros mostró niveles de hasta el 17% (terneros desde el nacimiento hasta los 6 meses de vida) en las granjas lecheras. Las altas tasas de mortalidad y morbilidad previas al destete van en contra de la economía de la granja y además de la salud y el bienestar de los animales.
El rumen, el punto clave


El epitelio ruminal es responsable de varias funciones fisiológicas muy importantes: la absorción, el transporte, el metabolismo de los ácidos grasos de cadena corta, entre otras. Al contrario de los otros órganos, que disminuyen su función como porcentaje del “peso corporal vacío” a medida que el rumiante madura, el rumen aumenta del 30 al 70% de la capacidad del sistema gastrointestinal durante el proceso del destete (Figura 1).
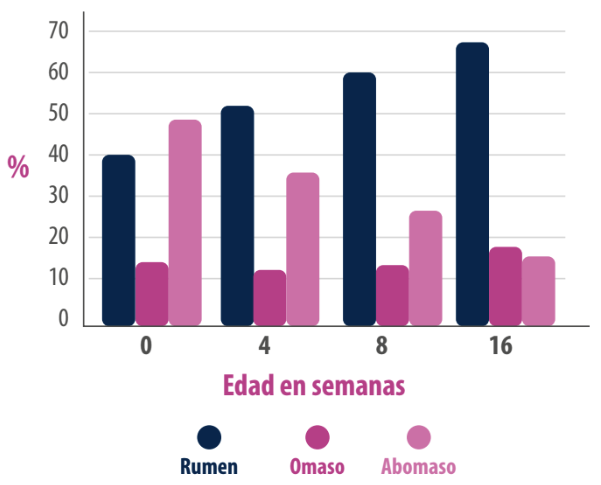

Figura 1. Desarrollo de los divertículos gástricos en terneros.


Es el hígado el órgano que controla los nutrientes disponibles que van hacia la periferia para sustentar el crecimiento, y debe adaptarse a los patrones de nutrientes absorbidos como resultado del desarrollo del rumen. En el pre-rumiante, en el hígado se lleva a cabo la glucólisis en un porcentaje muy alto, y la gluconeogénesis está, todavía, muy reducida en el total del metabolismo hepático.
![]()

Desarrollo y etapas
Al nacimiento el rumen está poco desarrollado tanto físicamente como metabólicamente, en el neonato no presenta el alto grado de queratinización característico del órgano maduro; metabólicamente el rumen, esencialmente no funciona con respecto a la capacidad cetogénica (desdoblamiento de los carbohidratos o fermentación).
Tras el inicio de la ingesta de alimento sólido y el posterior establecimiento de la fermentación ruminal, empieza su desarrollo físico y metabólico, el físico se puede dividir en dos:
1) Aumento de la masa ruminal (el grosor de la pared)
2) Crecimiento de las papilas
Las primeras investigaciones indicaron que la estimulación física por el alimento en el rumen podría explicar aumentos medibles, tanto en el peso del rumen, como en la musculatura.
![]()

Para que el epitelio ruminal avance, se debe establecer un proceso de fermentación viable y constante, lo que sugiere que se necesita la presencia de ácidos grasos de cadena corta o ácidos grasos volátiles (AGV) en la luz ruminal para promover el desarrollo papilar normal.
Los animales alimentados con grano y heno tienen un desarrollo armónico del rumen tanto del peso como de la capacidad de absorción, por el contrario, los neonatos alimentados únicamente con leche durante los primeros meses de vida tienen un desarrollo ruminal limitado con respecto: al peso ruminal, la capacidad, crecimiento papilar, el grado de queratinización, la pigmentación y el desarrollo de la musculatura.
Muy probablemente la falta de desarrollo ruminal se deba a la desviación efectiva de la leche directamente al abomaso mediante el cierre reflejo de la gotera esofágica, lo que impide que la leche o el sustituto lácteo ingrese al rumen provocando fermentación.
![]()

Lane y Jesse (1997) realizaron una infusión del 50% de las necesidades energéticas netas en corderos en forma de AGV a concentraciones fisiológicas que dio como resultado un aumento en la longitud de las papilas. Las infusiones de propionato de sodio y butirato de sodio, dieron lugar a un marcado desarrollo de las papilas ruminales en los terneros.
Una mezcla de sales de AGV (propionato y butirato incluidos) suplementadas dentro del 10% de una ración inicial de concentrado, dio lugar a un aumento de la incidencia de paraqueratosis ruminal, y en todos los animales tratados se observó un engrosamiento del estrato córneo tanto en terneros como en corderos.
El aumento de las cantidades de concentrado en la dieta no produjo ningún cambio en la musculatura del rumen, pero sí dio lugar a un aumento en la densidad y altura de las papilas en terneros y corderos.
No se ha podido identificar un único mecanismo responsable del desarrollo papilar. Sin embargo, existen varios procesos como: el metabolismo del butirato y el propionato en el epitelio ruminal que provoca un aumento del flujo sanguíneo a través del rumen, o el efecto directo del butirato o el propionato sobre la expresión génica dentro del rumen.
![]()

Estos genes tienen un patrón de expresión diferencial a lo largo del desarrollo de los animales criados normalmente, la expresión aumentó gradualmente, lo que indica un control más ontogénico* en lugar de un cambio dado por un desencadenante externo, en la expresión de estos productos génicos.


(*: «La ontogenia es la historia del cambio estructural de una unidad sin que ésta pierda su organización. Este continuo cambio estructural se da en la unidad, en cada momento, o como un cambio desencadenado por interacciones provenientes del medio donde se encuentre o como resultado de su dinámica interna»)
Un aumento rápido, pero no sostenido de butirato en el rumen, que no sucede fisiológicamente, estimula la proliferación celular, como lo indican los mayores índices mitóticos.


Dado que los ensayos in vitro dan resultados totalmente contrarios a los resultados in vivo, las diferentes respuestas in vivo e in vitro, y la naturaleza aparentemente contradictoria de los informes in vivo, todo esto sugiere una vía indirecta de estimulación celular. Se ha demostrado que los índices mitóticos epiteliales ruminales son estimulados por infusiones intravenosas de insulina.
Por otro lado, y debido a que se ha demostrado que el propionato es un estimulador de la liberación de insulina in vivo, es posible que la insulina pueda ser un mediador en la estimulación de la mitosis en el epitelio ruminal. Por lo tanto, otros factores que no sean la acción directa de los nutrientes no pueden ser eliminados como posibles agentes que controlan la proliferación epitelial ruminal, aunque aún no se han encontrado de manera concluyente.


Los ácidos grasos y la glucosa absorbidos en el intestino delgado deben pasar primero por el hígado; por lo tanto, la glucosa es el sustrato energético primario del tejido inmaduro, como es el caso de otros tejidos neonatales.
La absorción de oxígeno por el rumen neonatal es mayor cuando la glucosa está presente como sustrato oxidable en la luz ruminal; sin embargo, también el consumo de oxígeno por parte de las papilas ruminales maduras aumentó por encima de la absorción basal de oxígeno cuando se añadió glucosa.
Por el contrario, la adición de butirato estimuló la absorción de oxígeno en mayor grado en las papilas ruminales maduras que en el rumen neonatal. La cetogénesis del butirato también fue sustancialmente menor en el rumen neonatal en comparación con el tejido maduro, lo que podría indicar que, en este momento de la vida del animal, el butirato se usa directamente en el tejido epitelial del rumen.
Lane et al. (2000) evaluaron el desarrollo metabólico del epitelio ruminal en ausencia de ingesta de alimento sólido, manteniendo corderos únicamente con sustituto de leche y observaron el aumento característico y marcado en la capacidad cetogénica a los 42 días, independientemente del régimen dietético.
Si bien otros parámetros metabólicos no siguieron este desarrollo característico, esto es nuevamente indicativo de una respuesta ontogénica en lugar de un evento desencadenado por nutrientes.
Además, utilizando el ARN epitelial del rumen aislado de estos mismos animales de experimentación, demostró un aumento en las transcripciones génicas, a pesar de la falta de una producción significativa de AGV en el rumen.


La segunda parte de este contenido se publicará en la edición noviembre de nutriNews España
Te puede interesar Qué es la fibra, cómo cuantificarla y por qué