


La salmonella es un problema insidioso en la industria avícola significando un peligro crítico para la inocuidad de los alimentos. Además, también hay ya informes que confirman que serovares no tifoideos de salmonella causan salmonelosis en pollos (Ogunleye y Carlson 2012, Gong et al., 2016).
Por lo tanto, es de suma importancia identificar las estrategias de mitigación para la industria avícola.
La mayoría de Salmonella, excepto los serovares S.Pullorum y S.Gallinarum (Wilson et al., 2000) y posiblemente otras cepas (por ejemplo, S. Kentucky (Ogunleye y Carlson 2012), son capaces de residir asintomáticamente en los tractos intestinales de aves de corral.

Muchos serotipos de Salmonella pueden ser transmitidos por vía fecal-oral y ser luego excretados con las heces (Traub-Dargatz et al., 2006).
De este modo, la Salmonella es omnipresente en el ambiente, y por lo tanto, puede contaminar la carne de aves de corral antes del excretado fecal o durante el procesamiento de la canal (por fugas intestinales), resultando en una de las principales causas de infecciones por Salmonella en humanos (CDC, 2015).
A su vez, la resistencia a los antibióticos también se comprobó y se demostró que los metabolitos de fermentación de SC* pueden modular el microbioma (Kirkup y Riley, 2004), alterando los antibiogramas de las bacterias residentes (Mullins et al., 2013; Price et al., 2010).
Estudios recientes han demostrado que los metabolitos de fermentación de metabolitos de fermentación de SC* mejoran la salud y el rendimiento de las aves de corral (Jensen et al., 2008) al promover funciones inmunitarias tales como inducir la producción de anticuerpos antivirales, aumentar la actividad de lisozima en suero y aumentar la producción de IgM, linfocitos T y IgA secretora (Gao et al., 2008; Gao et al., 2009).
Possemiers et al., 2013, demostró ya en pruebas in vitro que los metabolitos de fermentación de SC* eran capaces de aumentar la producción de butirato intestinal; ácido graso de cadena corta que inhibe los mecanismos de virulencia de salmonella por la regulación de 17 genes (incluyendo hilA) en el “pool” de patogenicidad de salmonella (Gantois et al., 2006).
Esta represión en hilA fue más evidente en S.Enteriditis donde se observó una disminución de 6 veces en la expresión a través de un mecanismo desconocido (Gantois et al., 2006).
- Específicamente, los metabolitos de fermentación de SC* propiciaron una reducción significativa tanto en el número como en la prevalencia de Salmonella Heidelberg en pollos de engorde (Hofacre et al., 2015).
- Estos efectos anti-salmonella (disminución de la carga y prevalencia) también se han observado en terneros alimentados con metabolitos de fermentación de SC* donde el producto también protegió al animal de diversas consecuencias fisiopatológicas del patógeno (Brewer et al., 2014).
- Debido a estos posibles beneficios de seguridad alimentaria, se diseñó un estudio para examinar más detalladamente los efectos anti- Salmonella Typhimurium asociados con cultivo de levaduras en pollos.
OBJETIVO
El objetivo de este estudio fueron determinar los efectos de los metabolitos de fermentación de SC* sobre una Salmonella Typhimurium multi-resistente a antibióticos en pollos de engorde infectados experimentalmente con ese microbio.

MATERIAL & MÉTODOS
- Se llevó a cabo un experimento en la Universidad Estatal de Iowa (Ames) utilizando pollos de engorde Cobb de un día de edad.
- Tres repeticiones separadas e independientes de este experimento utilizando un total de 50 pollitos por experimento (25 por grupo de tratamiento), dando como resultado un total de 75 pollitos por tratamiento.
- Todas las aves fueron alimentadas con un pienso iniciador no medicado (con 24% PB) desde d0 a d21.
Todas las aves fueron pesadas individualmente en D21 y luego nuevamente al final del estudio en D49.
En tres ocasiones entre el d1 y Dd20, los pollos de engorde fueron infectados experimentalmente con múltiples Salmonella Typhimurium resistente a los antibióticos.
Después de confirmar que existía diseminación fecal de Salmonella en d21, los pollos de engorde fueron alimentados con una dieta que contenía metabolitos de fermentación de SC* (n = 57 aves, 1.25 kg /t pienso) o una dieta de control libre de metabolitos de fermentación de SC* (CON = 57 aves) hasta el d49.
Se obtuvieron muestras fecales semanalmente y se sometieron a cultivo selectivo para enumerar y determinar la resistencia antibiótica de Salmonella.

Tratamientos
- Sólo pienso de control de acabado (CON)
- Pienso de acabado que contenía 1,25 kg/t metabolitos de fermentación de SC*
Del d21 al d49, la dieta basal fue una dieta de acabado sin medicar (con 18-19%PB) con acceso ad libitum a comida y agua
A continuación, los aislados de Salmonella se sometieron a un ensayo de virulencia in vitro, que predice la capacidad de Salmonella para causar enfermedad en un huésped mamífero.
Los pollos de engorde se sacrificaron el d49 y se retiró un segmento del intestino grueso que se sometió a los mismos ensayos utilizados para las muestras fecales.
El protocolo usado en estas pruebas fueron aprobados por el Institutional Animal Care and Use Committee at Iowa State University.
Los métodos de evaluación y estadísticas de los resultados se pueden consultar en el texto original en el que se basa este artículo.
RESULTADOS
En todas las evaluaciones :
CON = Dieta control – sin metabolitos de fermentación de SC*
T = con metabolitos de fermentación de SC* (1,25 kg/t pienso)
1/ EVALUACIÓN DE LA DISEMINACIÓN FECAL DE SALMONELLA
Aves infestadas oralmente con Salmonella Typhimurium con multirresistencia antibiótica los d2, d9 y d16. aislaron.
Se aisló Salmonella a partir de heces (usando agar XLD) los d21,d28 y d42., y se inocularon Salmonella Typhimurium resistentes a antibióticos en D2, en D21, 28, 35 y 42.
Los datos representan la media ± SEM para 3 experimentos fueron realizados por separado. A, bP <0,01, a,c0.01<P<0.05
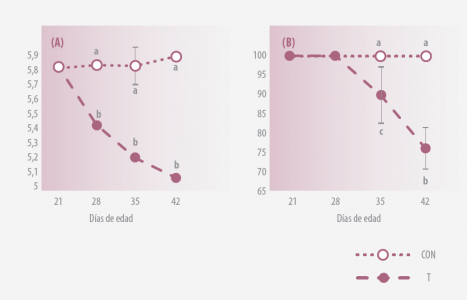

Figura 1. (A) Diseminación fecal de Salmonella* (UFC/g) (B) Prevalencia de diseminación fecal de Salmonella (% de aves)
2/ EVALUACIÓN DE LA VIRULENCIA DE SALMONELLA RECUPERADA DE LOS POLLOS EN PRUEBA
Todas las aves fueron sacrificadas el d49 y se extrajo una sección del intestino grueso y se cultivaron selectiva.
Los datos representan la media ± SEM para los 3 experimentos realizados por separado. A, bP < 0,01.
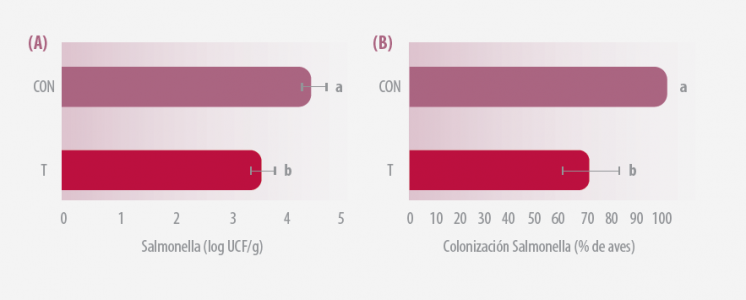

Figura 2. (A) Colonización de intestino grueso por Salmonella en d49 (B) Prevalencia de la colonización intestinal por Salmonella el d49
3/ EVALUACIÓN DE SU INVASIÓN TISULAR Y SU EXPRESIÓN GÉNICA EN HECES
Aproximadamente el 20% de la Salmonella recuperada de las aves se sometió a una RT-PCR semi-cuantitativo que evalúa la expresión de hilA (Carlson et al., 2007), el regulador global de la invasión de Salmonella (Bajaj et al., 1995).
El ARN se aisló y se sometió al ensayo semi-cuantitativo de RT-PCR en el que se documentó el número de ciclos de PCR (5 a 40) necesarios para visualizar un amplicón en electroforesis en gel de agarosa (Carlson et al., 2007).
El análisis de genes y la expresión se calcularon como porcentaje de CON, es decir, 100 x (número mínimo de ciclos requerido para visualizar un amplicón por muestras CON / número mínimo de ciclos requerido para visualizar y amplicon para muestras T).
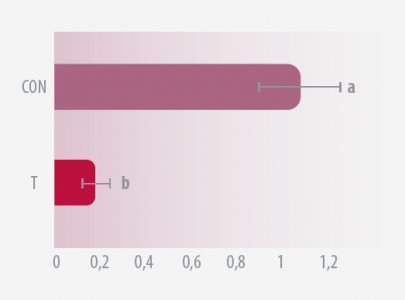

Figura 3a. Invasión tisular de Salmonella de pollos desafiados con Salmonella Typhimurium Figura
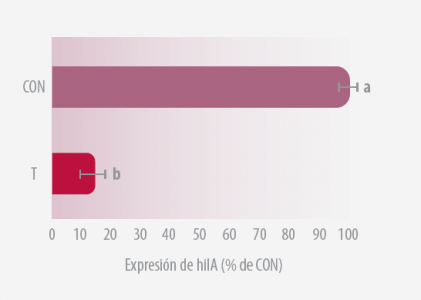

Figura 3b. Expresión hilA en Salmonella recolectada de las heces de los pollos desafiados con Salmonella Typhimurium
4/ EVALUACIÓN DE LA RESISTENCIA ANTIBIÓTICA DE LA SALMONELLA RECUPERADA DE LOS POLLOS EN PRUEBA
Las colonias de Salmonella recuperadas se inocularon individualmente en medio LB en placas de 96 pocillos en ausencia de cloranfenicol.
El crecimiento bacteriano se sometió entonces a un ensayo de PCR desarrollado y descrito previamente por Carlson et al. (1999).
El porcentaje de SGI1 (+) se calculó como 100 x (número de pocillos en los que Salmonella se replicó como amplicón específico SGI1 / 96).
Los datos representan la media ± SEM para los 3 experimentos realizados por separado. A, bP <0,01, a, c0,01 <P <0,05
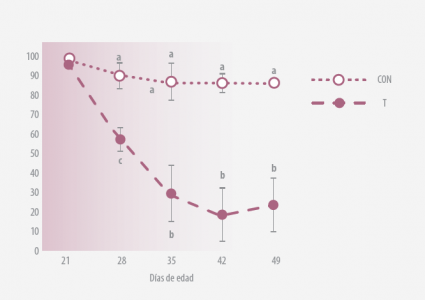

Figura 4. Presencia de SGI1 en Salmonella recuperada de las heces (días 21, 28, 35 y 42) o de intestinos (d49) de pollos de engorde infestados con Salmonella Typhimurium
En comparación con las aves alimentadas con la dieta CON,la diseminación fecal de Salmonella, la virulencia (invasión e invasión de expresión génica) y la resistencia a antibióticos disminuyeron significativamente en aves alimentadas con metabolitos de fermentación de SC* (5 veces, 7,5 veces, 6 veces y 5,3 veces de disminución, respectivamente).


El uso en inicio de metabolitos de fermentación de SC* redujo la virulencia y la resistencia a antibióticos de la cepa de Salmonella, y sus efectos aunque variables son beneficiosos permitiendo obtener un efecto positivo marcado en la seguridad alimentaria de la industria avícola.
Este estudio reveló que el metabolitos de fermentación de SC* inhibe la diseminación, la virulencia y la resistencia a los antibióticos de Salmonella que residen en pollos de engorde.
(*) Los metabolitos de fermentación de SC referenciados en el artículo corresponden a Diamond V Original XPCTM