")



In this article we will talk about the metabolism of volatile fatty acids (VFA) or short-chain fatty acids (SCFA).
RUMINAL DIGESTION
In the rumen environment or ecosystem, energy and protein are generated.
More than 60% of the energy that the animal uses for maintenance and production (growth, lactation, gestation, wool, etc.) comes from volatile fatty acids (VFA), and between 60 and 80% of the protein necessary for growth and production comes from the microbial population of the reticulum-rumen.
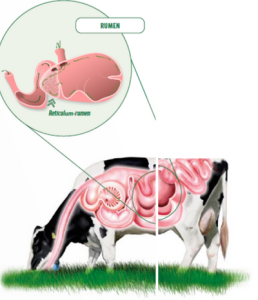
Therefore, the degradation and absorption of different feed components at the rumen level will determine future animal production (milk, meat, or wool).
Thus, the rumen represents a key segment of the digestive tract when it comes to ensuring optimal productive performance as well as to withstand extreme conditions.
Hence, if feed digestion within the rumen is enhanced, animal production will substantially improve and with it, the productivity of livestock systems as a whole.
FIBROUS CARBOHYDRATES
All foods (fodder or concentrates) are made up of different fractions: carbohydrates, proteins, lipids, vitamins, minerals and water. From these different food fractions, carbohydrates represent the most important energy source and the main precursors for the fat and sugar (lactose)found in milk.
Rumen microorganisms enable the host to obtain energy from fibrous carbohydrates (cellulose and hemicellulose):
Fiber in general, tends to be bulky and is retained in the rumen where it ferments slowly. Serving as a support for rumen microorganisms (MOOs).
There are different fiber qualities depending on the type and place of growth of the plant as well as its age.
As the plant matures, lignin content in fiber increases while the degree of ruminal fermentation of both cellulose and hemicellulose is reduced.
Lignification in plants does not only occur due to aging as it also depends on climate. For example, under increased heat conditions lignification also increases. The presence of fiber in long particles is necessary to stimulate rumination.
Rumination increases fiber separation and fermentation, stimulates rumen contractions, and increases the flow of saliva into the rumen.
Saliva contains bicarbonate and phosphates which contribute to maintaining the acidity (pH) of rumen content and pH values close to neutrality.
NON-FIBROUS CARBOHYDRATES
Non-fibrous carbohydrates (starches and sugars) ferment rapidly and in a large percentage within the rumen.
The presence of non-fibrous carbohydrates increases dietary energy density, improves energy supply as well as determining the amount of bacterial protein which is produced in the rumen.
However, non-fibrous carbohydrates do not stimulate rumination nor saliva production and when found in excess they can inhibit fiber fermentation.
![]() Thus, an adequate balance between fibrous and non-fibrous carbohydrates is very important in ruminant feeding in order to obtain profitable production levels.
Thus, an adequate balance between fibrous and non-fibrous carbohydrates is very important in ruminant feeding in order to obtain profitable production levels.
RUMEN FERMENTATION
PROPIONATE
The rumen digestion of starch generates a high production of VFA. Highlighting the production of propionate (C3) whose molar proportion increases in regards to the rumen fermentation of fibrous forage. Generating a higher molar proportion of acetate (C2).
Propionate found in the rumen is absorbed through the walls (more than 80%) reaching the liver. It is then transformed into glucose through the metabolic process known as gluconeogenesis.
ACETATE AND BUTYRATE
Meanwhile, acetate and butyrate directly generate energy in the form of ATP in the rumen (Van Huotert, 1993).
In high-producing animals, the main organs involved in carbohydrate metabolism are: the rumen, liver, and mammary gland.
CARBOHYDRATES
During rumen fermentation, microorganism populations, mainly bacteria, ferment carbohydrates to produce energy, gases (methane – CH₄ and carbon dioxide – CO₂), heat and organic acids.
The pathways involved in the formation of final fermentation products are very complex. Involving different reactions, that lead from pyruvate or phosphoenolpyruvate (glycolysis products), to CO₂, CH₄ and acetic, propionic, butyric and valeric acids (VFA).
ACETATE AND CO2
Acetate and CO₂ are produced by numerous species of rumen microorganisms from carbohydrates, while other species produce them from propionic, butyric and valerianic acids.
However, many of these individual species also have one or more end products such as succinate, lactate, formate, hydrogen, and ethanol, which are not final products of fermentation.
These compounds, except for ethanol, are subsequently metabolized by other species which produce the typical end products.
VFA
VFAs are end products of microbial fermentation which are absorbed through the keratinized epithelium of the rumen.
These acids are short chain carbon compounds, which are produced during the fermentative degradation of food and can be converted into glucose, amino acids or fatty acids by rumen bacteria or by the animal’s cells.
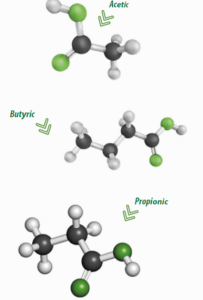
Formic, acetic, propionic, butyric , isobutyric, 2-methyl butyric, valeric, isovaleric, caproic and caprylic acids are considered VFAs.
Acetic, propionic and butyric acids are the ones produced in greater quantities. These are produced by intra- and extracellular enzymes of bacteria and protozoa.
Most of the acetate and all of the propionate are transported to the liver, On the other hand, most of the butyrate is converted to a ketone known as [beta]-hydroxybutyrate in the rumen wall. (Figure 1)
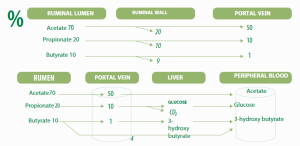
Figure 1. Absorption and metabolism of volatile fatty acids
Acetic, propionic and butyric acids make up the majority (>95%) of the acids produced in the rumen.
The fermentation of amino acids in the rumen also produces acids, which are called iso-acids.
The energy and iso-acids which are produced during fermentation are used by bacteria to grow (i.e. mainly to synthesize protein).
![]()
Ethanol, succinate, and lactate are catabolized to final products: acetic, propionic and butyric by the different microorganism species(Santini, 1994).
Acetic acid is transported in the bloodstream from the rumen and incorporated directly into the cellular proto-plasma. It is then used for obtaining energy via the acetyl-CoA and Krebs cycle, or for fat sinthesis(both fat tissue and milk fat).
CO₂ and CH₄ are burped, and the energy which is still present in CH₄ is lost.
The heat produced during fermentation is dissipated, if it is not necessary for maintaining body temperature.
The degradation and fermentation of polysaccharides (complex carbohydrates) in the rumen occurs essentially in three steps:
01. Fixation
Attachment of microorganisms to forage particles, causing the dissociation of carbohydrates from the structure of the cell matrix.
02 Hydrolysis
Hydrolysis of released polysaccharides to monosaccharides.
03 Fermentation
Intracellular fermentation of monosaccharides, from which VFAs are obtained. Pyruvate and phosphoenolpyruvate are the main products obtained from intracellular fermentation of hexoses and pentoses. Microorganisms transform these into different end products of rumen fermentation through different metabolic pathways (Leng 1973).

KETONES
Ketones are the main energy source for most body tissues.
Ketones are mainly derived from butyrate production in the rumen, but they can also come from the mobilization of adipose tissues during the initial stages of lactation .
The liver is the main organ where the formation of ketone bodies occurs, while kidneys participate to a lesser extent.
The process takes place in the mitochondrial matrix and occurs in different steps.
CATABOLISM AND ANABOLISM

The nutrients absorbed in the rumen or small intestine, are mainly directed to the liver and other organs for catabolism or anabolism, as needed.
The metabolism of monosaccharides revolves around the supply and destination of hexose, this monomer being the main source of energy for cells, the catabolic pathway is glycolysis, carried out in the cell cytoplasm. This process consists of eight reactions:
- Glucose enters the cytoplasm to be phosphorylated (addition of a phosphate group), from ATP. This reaction is catalyzed by hexokinase. The resulting glucose 6-phosphate (aldohexose) is abundant in all cells, since the vast majority that enters the cytoplasm ends up being phosphorylated, in order to prevent it from crossing back the cytoplasmic membrane and diffusing to the extracellular environment.
- Aldohexose is isomerized and transformed into fructose 6-phosphate (ketohexose). Reaction catalyzed by glucose-6-phosphate isomerase.
- Fructose-6-phosphate is phosphorylated from ATP at carbons 1 and 6 to give rise to fructose-1,6 bisphosphate. Phosphofructokinase-catalyzed reaction.
- Fructose-1,6-bisphosphate is divided into two: glyceraldehyde-3-phosphate and dihydroxyacetone phosphate. Aldose-catalyzed reaction.
- Triose phosphate isomerase catalyzes the conversion of dihydroxyacetone phosphate to obtain more glyceraldehyde-3-phosphate
- Glyceraldehyde-3-phosphate is oxidized and phosphorylated, at carbons 1 and 6 forming 1,3-bisphosphoglycerate by glyceraldehydephosphate dehydrogenase, then transfers its phosphate group, to synthesize ATP and is transformed into 3-phosphoglycerate.
- 3-phosphoglycerate isomerizes C3 to C2 and is transformed into 2-phosphoglycerate by phosphoglycerate mutase. Subsequently, the enolase promotes the formation of a double bond, eliminating a molecule of H₂O and forming phosphoenolpyruvate.
- Phosphoenolpyruvate transfers its phosphate group to synthesize ATP and is transformed into pyruvate.
Pyruvate enters the mitochondrial matrix, therefore, each hexose (glucose) generates:
2 pyruvates, 2 ATPs, 2 NADH+H+, 2 hydrogen ions and 2 H₂O molecules, pyruvates are metabolized to acetyl-Coenzyme A, thus allowing their entry into the Krebs cycle and producing 12 ATPs
GLUCONEOGENESIS
All propionate is converted to glucose in the liver. In addition, the liver uses amino acids for glucose synthesis, this process is known as gluconeogenesis.
It is very important, because in general there is not enough glucose absorbed in the digestive tract and all the sugars found in milk (lactose approximately 1575g in a cow that produces 35 kg of milk a day) must be produced by the liver in forage diets.
Gluconeogenesis also synthesizes GLUCOSE from lactate and glycerol, in the cell cytoplasm and mitochondria of hepatocytes.
In this metabolic pathway, six ATPs are consumed for each hexose produced and the CH3-CH2-COOH propionate is the only glucogenic VFA.

The importance of glycogenesis in ruminants is mainly due to the fact that their body absorbs small amounts of glucose in the digestive tract and that its ability to store glycogen in the liver is more limited than in monogastric livers.






