



La formación de metano en animales rumiantes y estrategias de mitigación basadas en la alimentación. Parte I
El metano y el rumiante
Como ya se ha revisado en otros trabajos, el metano (CH4) entérico producido por los animales rumiantes es responsable aproximadamente del 6% de las emisiones antropogénicas globales de gases de efecto invernadero.
El CH4 tiene una vida útil mucho más corta (vida media; 8,6 años; Muller y Muller, 2017) que el CO2 en la atmósfera, lo que lo convierte en un objetivo de mejora a corto plazo en la reducción del calentamiento global.
| Las emisiones de CH4 por unidad de carne y leche han disminuido en los últimos años debido a mejoras en la eficiencia en la producción y rendimiento animal. |
- Las condiciones anaeróbicas del rumen deben mantener una armonía entre los microorganismos y los procesos de fermentación.
- El rumen es un ecosistema abierto y continuo en donde se permite la entrada de sustratos y la salida de productos.
- La formación de CH4 es un proceso fisiológico normal que contribuye al correcto funcionamiento fermentativo del rumen, contrarrestando la inhibición de otros microorganismos provocado por la acumulación de H2.
Arqueas metanógenas, los microorganismos que crecen en ausencia de oxígeno y producen CH4
Las arqueas (Tercer Reino Archaea, descubiertas en 1977 en la Universidad de Illinois por Wolfe R. y Woese C., y que le acuñaron el término arcaico) son microorganismos que crecen en ausencia de oxígeno y producen CH4.
Son un grupo de microorganismos unicelulares, similares a las bacterias (arqueobacterias: término mal empleado) en tamaño y forma, con morfología procariota (sin núcleo y orgánulos membranosos internos).
Las arqueas metanógenas se clasifican en cinco órdenes:
- Methanobacteriales
- Methanococcales
- Methanomicrobiales
- Methanosarcinales
- Methanopyrales
La mayoría de las especies son inmóviles, sólo los miembros del género Methanothermus se desplazan por un flagelo.
↦ Dependiendo del sustrato utilizado, los metanógenos se clasifican en tres grupos:
- hidrogenótrofos (los más abundantes, utilizan los iones H2 para reducir el CO2)
- metilótrofos (utilizan compuestos metílicos)
- acetótrofos (utilizan el grupo metilo del acetato)


Termodinámica del rumen
Los carbohidratos son la principal fuente de energía de la dieta de los rumiantes. Los carbohidratos ingeridos por los rumiantes se degradan a monómeros por acción microbiana.
Los diferentes microorganismos del rumen utilizan energía para mantener su organización, crecimiento y reproducción.
⇒ Energía libre de Gibbs (DG)
Los procesos de fermentación en el rumen se pueden estudiar conociendo la variación de la energía libre de Gibbs (DG).
La energía libre de Gibbs es la parte de la energía de un sistema que se puede convertir libremente en trabajo útil a temperatura y presión constante.


La fórmula está formada por:
- la entalpía (DH) definida como el contenido energético del sistema (por ejemplo, energía almacenada en los enlaces químicos)
- la entropía (DS) que mide la parte no utilizable de la energía contenida en un sistema.
- T es la temperatura que multiplica a la entropía y se expresa en grados Kelvin.
El cambio en la energía libre de Gibbs durante una reacción metabólica nos va a dar información sobre la espontaneidad de ese proceso (es decir, si puede llevarse a cabo sin aporte externo de energía).
- Si el valor de DG es positivo, la reacción no es espontánea y no puede tener lugar sin un aporte de energía exterior.
- Si el valor de DG es negativo, se libera energía y la reacción es espontánea generando un producto.
Es decir, la energía libre Gibbs en un sistema abierto como es el rumen mide los cambios que se producen entre un estado inicial y otro final:
- energía liberada (DG negativo) o
- energía absorbida (DG positivo)
Parte de la energía negativa libre de Gibbs, asociada a los procesos fermentativos, es utilizada por los microorganismos del rumen para generar ATP que puede ser utilizado para el crecimiento microbiano, transporte activo movilidad, etc.

⇒ Además, el crecimiento microbiano produce proteína microbiana, que es la principal y más económica fuente de aminoácidos para los rumiantes.
Uno de los productos de la fermentación microbiana es el CH4; los metanógenos ruminales utilizan los electrones del H2 y reducen el CO2 a CH4 en secuencias acopladas a la síntesis de ATP, donde el CO2 es la fuente de carbono y el H2 es el principal donador de electrones.
| Temperatura y presión constantes de 25º (298ºK) y 1 atm, respectivamente y siguiendo la fórmula anterior (DG= DH – TDS) la molécula de CH4 tiene una variación en la energía libre de DG = -68 kJ/mol, es decir, se trata de una reacción que sucede de forma espontánea. |
⇒ Los carbohidratos de los alimentos poseen en sus enlaces una elevada cantidad de energía libre de formación.


⇒ Esta energía se va liberando en diferentes rutas fermentativas en el rumen por acción de los microorganismos.
Es decir, el gran número de rutas y reacciones que puede seguir la glucosa en el rumen está parcialmente controlada por los productos finales que se generan durante la fermentación.
La Figura 1 muestra algunos datos termodinámicos (energía libre de formación en condiciones estándar) de diversos reactantes y productos del metabolismo ruminal, junto a otros importantes metabolitos ruminales; por ejemplo:
- acetato; DG = -318 kJ/mol
- butirato; DG = -312 kJ/mol
- propionato; DG = -322 kJ/mol
- lactato; DG = -517 kJ/mol
| Cuando la DG en una reacción está cercana a cero (equilibrio), hay poca energía disponible que puedan utilizar los microorganismos para el crecimiento. |
Es decir, cuando DG para la producción de CH4 es cercano a 0, el crecimiento de los microorganismos metanogénicos se ralentiza debido a la escasa disponibilidad H2.
⇒ Rutas captadoras de [H]
En el rumen, los polisacáridos (principalmente celulosa, hemicelulosa y almidón) se hidrolizan a glucosa y otras hexosas y pentosas.
↳ Los monosacáridos son además metabolizados a ácidos grasos volátiles (AGV) y CO2.
↳ El hidrógeno metabólico ([H]) se libera en el metabolismo de los monosacáridos hacia los AGV, reduciendo cofactores intracelulares, y para que la fermentación continúe, los cofactores deben ser re-oxidados.
La Figura 1 nos muestra diferentes rutas captadoras de [H] así como las variaciones en la energía libre de formación de Gibbs.
↪ Esto sucede en gran medida a través de la actividad de la hidrogenasa y la formación de dihidrógeno (H2, es decir, hidrógeno molecular).
|
El H2 ruminal existe en dos formas:
|
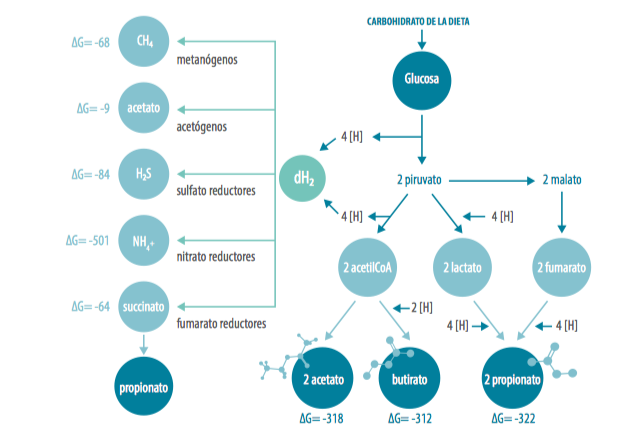

Figura 1. Esquema de principales rutas de fermentación en rumen incluyendo la generación e incorporación de H y H2. Estimación de las variaciones en la energía de Gibbs basadas en el trabajo de Ungerfeld y Kohn (2006) sin considerar la generación de ATP (ΔG en kJ/mol)
Solo dH2 está disponible para microorganismos ruminales (Wang et al., 2014).
↳ El dihidrógeno no se acumula en el rumen porque se transfiere desde el consorcio fermentativo formado por bacterias, protozoos y hongos hacia arqueas metanogénicas que lo utilizan para reducir el CO2 y otros compuestos de un carbono a través de la vía hidrogenotrófica hacia CH4.
- Formiato. La mayoría de los metanógenos también pueden utilizar el formiato generado en la formación de acetil-CoA a partir de piruvato, como donante [H] para la metanogénesis ruminal (Schauer y Ferry, 1980), convirtiéndose el formiato no utilizado en H2 y CO2.
- En menor medida, el CH4 también se puede producir en el rumen mediante la utilización de grupos metilo (ruta metilotrófica) y con menos frecuencia a partir de acetato (Huws et al., 2018).
|
El CH4 representa el sumidero más grande de [H] en el rumen. |
Es importante destacar que mientras que la mayoría de [H] producido en los procesos fermentativos por la microbiota se transfiere a los metanógenos como dH2 y se utiliza en la metanogénesis (Janssen, 2010),.
› Propionato. Hay otras importantes vías de utilización de [H] como la producción de propionato, que puede incorporar [H] en cofactores reducidos generado en reacciones intracelulares, así como dH2 producido por otras células (Henderson, 1980).
Una relación positiva entre las concentraciones de dH2 y propionato indica que el aumento de dH2 ruminal puede facilitar reacciones que incorporan [H] en la producción de propionato (Wang et al., 2016).
↳ El propionato es un sumidero [H] alternativo al CH4 (Janssen, 2010), principal precursor de la glucosa para los rumiantes y, por tanto, deseable de mejorar en animales con altas demandas de precursores glucogénicos.
› Acetato . La formación de acetato a partir de CO2 y H2 (acetogénesis reductora), también es un proceso de incorporación de [H] deseable, ya que el acetato es una fuente de energía y bloque de construcción en la síntesis de ácidos grasos de cadena larga.
Sin embargo, la acetogénesis reductora es termodinámicamente superada por la metanogénesis en el rumen (Ungerfeld y Kohn, 2006) [DG = -68 y -9 kJ/mol para la metanogénesis y acetogénesis reductora, respectivamente], pero sería un sumidero de [H] beneficioso para mejorar la fermentación ruminal cuando la metanogénesis está inhibida (Ungerfeld, 2013).
› Nitrato y sulfato . Algunos aceptores de electrones inorgánicos como el nitrato y el sulfato pueden competir superando termodinámicamente la metanogénesis en el rumen (Ungerfeld y Kohn, 2006), aunque la disponibilidad de estos [H] aceptadores es bajo con la mayoría de las dietas a menos que estos compuestos aceptadores de electrones se suplementen.
⇒ Redirigir [H] de la metanogénesis
En teoría, redirigir [H] de la metanogénesis hacia productos finales de la fermentación para que puedan ser utilizados por el animal huésped, así como la síntesis de biomasa microbiana, ayuda no solo a disminuir las emisiones de CH4, sino que pueden también potencialmente beneficiar la productividad del animal huésped.
Vamos a ver algunos ejemplos de re-dirección de los [H] mediante el manejo de la alimentación.
La manipulación dietética puede ser un camino de mitigación de CH4 muy eficaz, y hay muchas revisiones exhaustivas (Beauchemin et al., 2009; Hristov et al., 2013; Knapp et al., 2014; Beauchemin et al., 2020)).
La eficiencia de la mitigación de CH4 con una dieta en particular dependerá de:
- sus efectos sobre el flujo de H2 en el rumen y su concentración
- la comunidad microbiana
- vías de fermentación
- el tiempo de permanencia del alimento en el rumen y
- las interacciones entre estos factores.
En la segunda parte de este trabajo, se abordarán en detalle las Estrategias nutricionales de Mitigación de emisiones de CH4