




Importância da inclusão de taurina e L-carnitina na dieta de cães e gatos
 A suplementação de alguns nutrientes purificados em dietas destinadas a animais de produção é uma forma de simplificar as dietas as tornando completas e seguras do ponto de vista nutricional. Na alimentação de animais de companhia a prática também foi adotada e possibilita a formulação de dietas específicas para cães e para gatos.
A suplementação de alguns nutrientes purificados em dietas destinadas a animais de produção é uma forma de simplificar as dietas as tornando completas e seguras do ponto de vista nutricional. Na alimentação de animais de companhia a prática também foi adotada e possibilita a formulação de dietas específicas para cães e para gatos.
Assim, é comum que os tutores encontrem alimentos completos e balanceados para cães e gatos em que a taurina e a carnitina, dentre outras substâncias, estão elencadas na composição dos produtos.
| TAURINA |
 Cães, classificados como onívoros, e gatos, carnívoros estritos, possuem características metabólicas e necessidades nutricionais diferentes.
Cães, classificados como onívoros, e gatos, carnívoros estritos, possuem características metabólicas e necessidades nutricionais diferentes.
![]() Ao longo do processo de domesticação, os cães adquiriram capacidades metabólicas que os permitem sintetizar substâncias que os gatos não conseguem.
Ao longo do processo de domesticação, os cães adquiriram capacidades metabólicas que os permitem sintetizar substâncias que os gatos não conseguem.
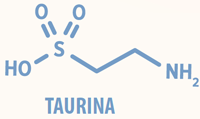
Exemplo é a taurina, um β-aminoácido sulfurado (ácido 2-aminoetanossulfônico) essencial para gatos, mas não-essencial para cães.
A estrutura química da taurina difere dos demais aminoácidos (Figura 1), o grupamento amino está ligado ao carbono β e não ao carbono α.
Adicionalmente, a taurina possui um grupamento sulfônico (-SO3) e não um grupamento carboxílico (-COOH). Assim, a taurina não se incorpora às proteínas permanecendo livre no citosol.
A biossíntese de taurina ocorre através dos aminoácidos sulfurados metionina e cisteína, que atuam como precursores. Através da atividade de duas enzimas chave, cisteína dioxigenase (CD) e cisteína sulfinato descarboxilase (CSD), o β-aminoácido é produzido endogenamente (Figura 1).
![]() Em cães, ambas as enzimas atuam eficientemente na síntese de taurina. No entanto, em gatos a CSD possui atividade reduzida no fígado, aproximadamente 200 a 300 vezes menor do que a atividade em ratos (Stipanuk, 1985), o que implica no desvio da cisteína para síntese de piruvato.
Em cães, ambas as enzimas atuam eficientemente na síntese de taurina. No entanto, em gatos a CSD possui atividade reduzida no fígado, aproximadamente 200 a 300 vezes menor do que a atividade em ratos (Stipanuk, 1985), o que implica no desvio da cisteína para síntese de piruvato.
 Adicionalmente, os gatos possuem reduzida atividade da CD e baixa utilização de cisteína como metabólito para síntese de taurina.
Adicionalmente, os gatos possuem reduzida atividade da CD e baixa utilização de cisteína como metabólito para síntese de taurina.
Em gatos, apenas 20% da cisteína disponível é utilizada para síntese de taurina, enquanto em ratos esse teor é de 80%. Desta forma, os gatos são dependentes da ingestão de taurina via dieta para suprir o requerimento, uma vez que suas vias metabólicas, a partir de aminoácidos sulfurados, é desviada para a produção de compostos que serão úteis na geração de energia ou gliconeogênese.
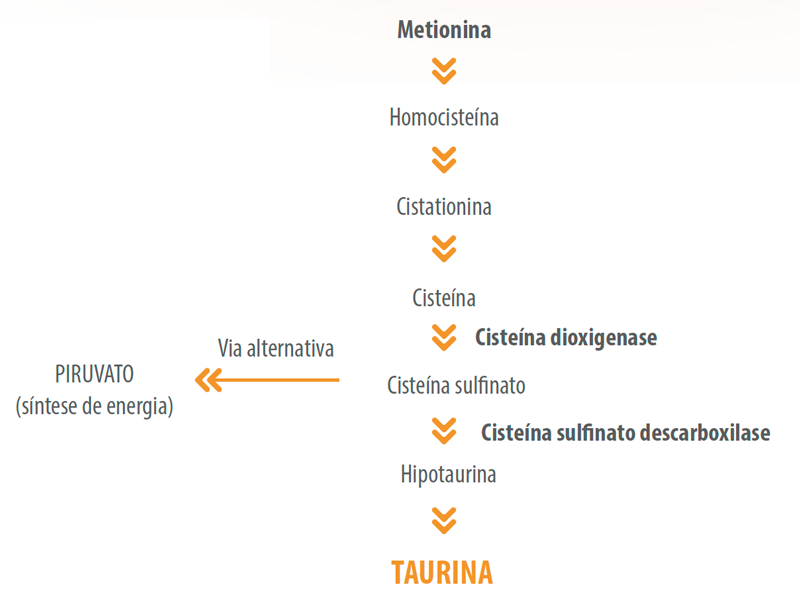
Figura 1. Estrutura química e síntese da taurina.

 Dentre as funções da taurina, destaca-se a sua participação na conjugação dos sais biliares que atuam na emulsificação dos lipídeos.
Dentre as funções da taurina, destaca-se a sua participação na conjugação dos sais biliares que atuam na emulsificação dos lipídeos.
Além da taurina, a glicina também pode ser conjugada, porém, os felinos não são capazes de utilizá-la sendo dependentes da taurina para a conjugação. A maior fração dos sais biliares são reabsorvidos pela circulação enterohepática no jejuno e íleo.
No entanto, alguns microrganismos são capazes de desconjugar os sais biliares, liberando a taurina que pode ser reabsorvida ou degradada por bactérias, resultando na sua perda.
A deficiência de taurina pode ser clinicamente constatada dentro de semanas ou meses, sendo a degeneração da retina e a cardiomiopatia dilatada os principais distúrbios associados à sua deficiência em cães e gatos, além de danos sobre as funções reprodutiva e neurológica.
Hayes et al. (1975) observaram a ocorrência de degeneração central da retina em gatos em menos de 6 semanas após alimentação com dietas purificadas isentas de taurina.
![]() A partir do exame de eletrorretinograma visualizou-se a redução da amplitude das ondas dos receptores do tipo cone.
A partir do exame de eletrorretinograma visualizou-se a redução da amplitude das ondas dos receptores do tipo cone.
Pion et al. (1998) constataram que apenas alguns cães alimentados com dietas livres de taurina apresentaram lesões hiperreflectivas bilaterais, quadro clínico similar à degeneração central da retina.
A taurina pode ser encontrada em elevada disponibilidade em frutos do mar e tecidos animais, como:
![]() cérebro
cérebro
![]() musculatura esquelética
musculatura esquelética
![]() retina
retina
![]() vísceras como coração, fígado e rins.
vísceras como coração, fígado e rins.
Alguns insetos e algas também são capazes de sintetizar taurina, como Mazaella spp., Porphyra spp. e Chondracanthus spp. (McCusker et al., 2014).
![]()
 A cardiomiopatia dilatada, associada a sintomas secundários como desidratação, letargia e hipotermia, foi primeiramente relacionada à deficiência de taurina em gatos por Pion et al. (1987). A taxa de sobrevivência de gatos com cardiomiopatia dilatada moderada a severa consumindo dieta suplementada com taurina é de 60%, enquanto gatos consumindo dieta sem taurina é de 20% (Pion et al., 1992a; Pion et al., 1992b).
A cardiomiopatia dilatada, associada a sintomas secundários como desidratação, letargia e hipotermia, foi primeiramente relacionada à deficiência de taurina em gatos por Pion et al. (1987). A taxa de sobrevivência de gatos com cardiomiopatia dilatada moderada a severa consumindo dieta suplementada com taurina é de 60%, enquanto gatos consumindo dieta sem taurina é de 20% (Pion et al., 1992a; Pion et al., 1992b).
 Os distúrbios ligados às funções reprodutiva e neurológica são mais brandos comparados aos reportados na retina e coração. Dentre os danos reprodutivos, existem registros de reabsorção uterina, abortos e natimortos (Sturman et al., 1985; Sturman et al., 1986). Além disso, algumas raças apresentam predisposição como os cães Newfoundland, em que baixos níveis plasmáticos e totais de taurina podem desencadear problemas reprodutivos.
Os distúrbios ligados às funções reprodutiva e neurológica são mais brandos comparados aos reportados na retina e coração. Dentre os danos reprodutivos, existem registros de reabsorção uterina, abortos e natimortos (Sturman et al., 1985; Sturman et al., 1986). Além disso, algumas raças apresentam predisposição como os cães Newfoundland, em que baixos níveis plasmáticos e totais de taurina podem desencadear problemas reprodutivos.
Os tecidos vegetais não possuem taurina, e bactérias possuem apenas pequenas quantidades. No entanto, fontes purificadas na forma cristalina estão disponíveis comercialmente podendo ser utilizadas para suplementação em dietas para cães e gatos.
De modo geral, grande parte das dietas completas e balanceadas para cães atualmente disponíveis no mercado não são suplementadas com taurina, pois os níveis dos precursores metionina e cisteína presentes na fórmula atendem aos requerimentos necessários para as suas funções e à quantidade a ser utilizada para a biossíntese de taurina.
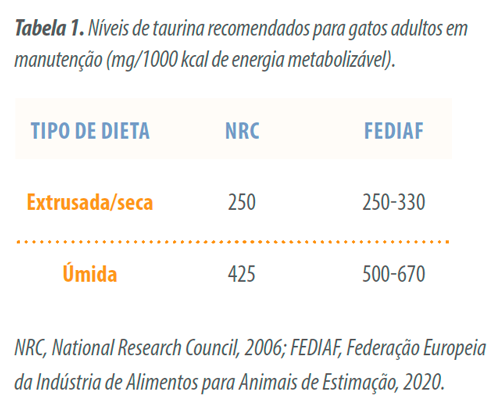
 Para gatos, a suplementação com taurina é prática comum em alimentos comerciais devido às características metabólicas intrínsecas da espécie previamente mencionadas. A recomendação de suplementação de taurina depende do processamento aplicado ao alimento (Tabela 1). Devido a sua elevada solubilidade em água, a taurina pode ser dissolvida quando exposta à umidade.
Para gatos, a suplementação com taurina é prática comum em alimentos comerciais devido às características metabólicas intrínsecas da espécie previamente mencionadas. A recomendação de suplementação de taurina depende do processamento aplicado ao alimento (Tabela 1). Devido a sua elevada solubilidade em água, a taurina pode ser dissolvida quando exposta à umidade.
Assim, dietas úmidas são mais propensas a perda de taurina caso a água de cocção seja removida e descartada. Já as dietas extrusadas ou assadas possuem maior retenção de taurina, porém com algumas perdas durante o processamento (Spitze et al., 2003).
| L-CARNITINA |
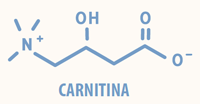 A carnitina, nutriente essencial condicional, é uma amina quaternária (3-hidroxi-4-N-trimetilamino-butirato) que pode ser sintetizada no fígado e rins a partir dos aminoácidos essenciais lisina e metionina (Figura 2). A biossíntese de carnitina demanda ainda a presença de ferro, ácido ascórbico, niacina e vitamina B6.
A carnitina, nutriente essencial condicional, é uma amina quaternária (3-hidroxi-4-N-trimetilamino-butirato) que pode ser sintetizada no fígado e rins a partir dos aminoácidos essenciais lisina e metionina (Figura 2). A biossíntese de carnitina demanda ainda a presença de ferro, ácido ascórbico, niacina e vitamina B6.
 Como acontece com a taurina, é comum que os tutores encontrem declarado na composição de alimentos completos e balanceados para cães e gatos a presença de carnitina. Especialmente em dietas light, sênior e terapêuticas, como aquelas dietas coadjuvantes no tratamento de cardio e hepatopatias.
Como acontece com a taurina, é comum que os tutores encontrem declarado na composição de alimentos completos e balanceados para cães e gatos a presença de carnitina. Especialmente em dietas light, sênior e terapêuticas, como aquelas dietas coadjuvantes no tratamento de cardio e hepatopatias.
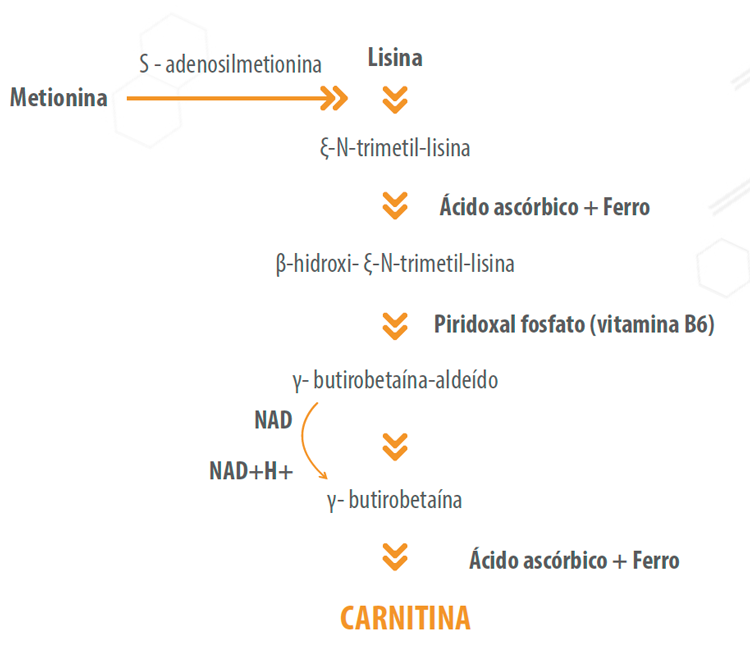
Figura 2. Estrutura química e síntese de carnitina.
 A carnitina está envolvida em passos importantes da produção de energia através da β-oxidação, na qual atua como transportador transmembrana de ácidos graxos livres de cadeia longa presentes no citosol celular para o interior das mitocôndrias (Kerner & Hoppel, 2000), permitindo assim a oxidação desses nutrientes e geração de adenosina trifosfato (ATP).
A carnitina está envolvida em passos importantes da produção de energia através da β-oxidação, na qual atua como transportador transmembrana de ácidos graxos livres de cadeia longa presentes no citosol celular para o interior das mitocôndrias (Kerner & Hoppel, 2000), permitindo assim a oxidação desses nutrientes e geração de adenosina trifosfato (ATP).
[registrados]
Devido ao seu tamanho, apenas os ácidos graxos de cadeia longa requerem carnitina para atravessar a membrana mitocondrial, uma vez que ácidos graxos de cadeia curta e média apresentam solubilidade suficiente para atravessar diretamente.
 O consumo de dietas contendo fontes proteicas com baixas concentrações de lisina, metionina ou ambas pode comprometer a biossíntese de carnitina. A diminuição da síntese endógena de carnitina promove a redução do transporte de ácidos graxos para o interior da mitocôndria e a sua oxidação é prejudicada, o que pode contribuir para o desenvolvimento de lipidose hepática, caracterizada como o acúmulo de lipídeos no fígado, em indivíduos em restrição energética exacerbada (Center et al., 2000).
O consumo de dietas contendo fontes proteicas com baixas concentrações de lisina, metionina ou ambas pode comprometer a biossíntese de carnitina. A diminuição da síntese endógena de carnitina promove a redução do transporte de ácidos graxos para o interior da mitocôndria e a sua oxidação é prejudicada, o que pode contribuir para o desenvolvimento de lipidose hepática, caracterizada como o acúmulo de lipídeos no fígado, em indivíduos em restrição energética exacerbada (Center et al., 2000).
Em humanos, a carnitina é amplamente utilizada por atletas como um agente termogênico devido à sua ação sobre a oxidação de ácidos graxos. Em animais de companhia, a sua utilização se concentra em alimentos destinados à animais em sobrepeso e obesidade, bem como para animais de trabalho ou que desenvolvam exercício prolongado.
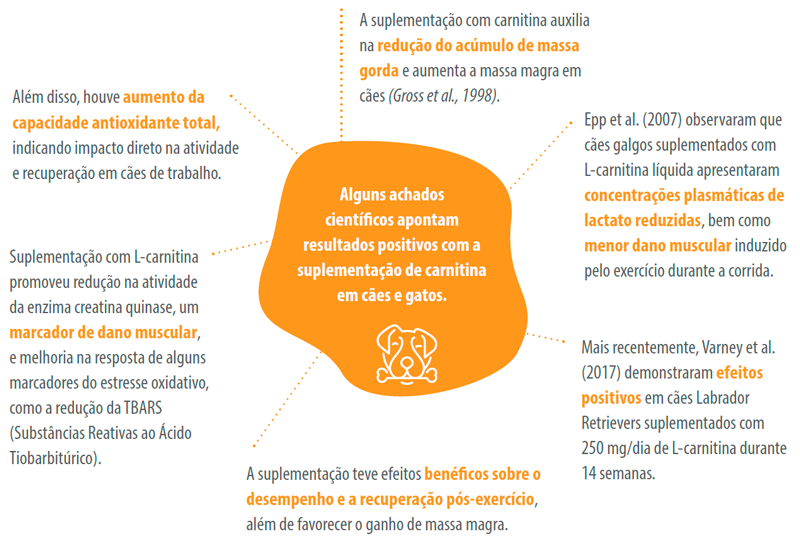

 Ao avaliar a suplementação com 1000 mg/kg de L-carnitina em gatos submetidos à lipidose hepática experimental, Blanchard et al. (2002) observaram o efeito protetor da L-carnitina no controle das concentrações plasmáticas de ácidos graxos e β-hidroxibutirato, marcadores da resposta ao jejum e/ou cetose. Em gatos com sobrepeso e submetidos a perda de peso rápida, a suplementação com L-carnitina promoveu aumento na taxa metabólica e na oxidação de ácidos graxos (Center et al., 2012).
Ao avaliar a suplementação com 1000 mg/kg de L-carnitina em gatos submetidos à lipidose hepática experimental, Blanchard et al. (2002) observaram o efeito protetor da L-carnitina no controle das concentrações plasmáticas de ácidos graxos e β-hidroxibutirato, marcadores da resposta ao jejum e/ou cetose. Em gatos com sobrepeso e submetidos a perda de peso rápida, a suplementação com L-carnitina promoveu aumento na taxa metabólica e na oxidação de ácidos graxos (Center et al., 2012).
Entretanto, inúmeros trabalhos científicos apontam resultados contraditórios quanto a eficiência da sua suplementação. Para cardiopatas, alguns achados são positivos (Keene et al., 1991) e em outros ausentes (Costa & Labuc, 1994).
![]() Em um relato de caso, Keene et al. (1991) verificaram que cães Doberman Pinschers com cardiomiopatia dilatada e deficiência de carnitina apresentaram melhora na saúde e na função cardíaca após a suplementação com 150 mg/kg/dia de L-carnitina via oral.
Em um relato de caso, Keene et al. (1991) verificaram que cães Doberman Pinschers com cardiomiopatia dilatada e deficiência de carnitina apresentaram melhora na saúde e na função cardíaca após a suplementação com 150 mg/kg/dia de L-carnitina via oral.
 A concentração orgânica de carnitina é resultado de processos metabólicos, como ingestão, biossíntese, transporte dentro e fora dos tecidos e excreção. Cães não catabolizam a L-carnitina (Yue & Fritz, 1962; Rebouche & Engel, 1983), de modo que a excreção urinária reflete o total absorvido e sintetizado. O consumo regular de L-carnitina associado ao acréscimo da gordura na dieta em cães promove o aumento na perda urinária de carnitina (Sanderson et al., 2001).
A concentração orgânica de carnitina é resultado de processos metabólicos, como ingestão, biossíntese, transporte dentro e fora dos tecidos e excreção. Cães não catabolizam a L-carnitina (Yue & Fritz, 1962; Rebouche & Engel, 1983), de modo que a excreção urinária reflete o total absorvido e sintetizado. O consumo regular de L-carnitina associado ao acréscimo da gordura na dieta em cães promove o aumento na perda urinária de carnitina (Sanderson et al., 2001).
Comercialmente, os alimentos secos light para cães e gatos possuem entre 140 e 500 mg/kg de L-carnitina na matéria seca. Por sua vez, as dietas terapêuticas possuem níveis mais elevados de suplementação, com valores entre 400 e 830 mg/kg de L-carnitina na matéria seca.
| TAURINA E L-CARNITINA, SUPLEMENTAR OU NÃO? |
Como visto, a suplementação com taurina em gatos é necessária frente a incapacidade enzimática e metabólica apresentada pela espécie. Assim, dietas comerciais completas e balanceadas destinadas a gatos, seja na forma extrusada ou úmida, obrigatoriamente devem possuir taurina na sua composição para permitir o balanço final dos nutrientes.
![]()
 Diferentemente, em cães a sua inclusão pode ser facultativa a depender das características químicas dos ingredientes utilizados na formulação da dieta.
Diferentemente, em cães a sua inclusão pode ser facultativa a depender das características químicas dos ingredientes utilizados na formulação da dieta.
![]() Quanto à suplementação com L-carnitina, não existem níveis de recomendação específicas para cães e gatos, assim a sua inclusão varia em dietas de manutenção, light e terapêuticas.
Quanto à suplementação com L-carnitina, não existem níveis de recomendação específicas para cães e gatos, assim a sua inclusão varia em dietas de manutenção, light e terapêuticas.
![]() Diante dos achados científicos contrastantes, a sua utilização em dietas para animais saudáveis ainda pode ser questionável.
Diante dos achados científicos contrastantes, a sua utilização em dietas para animais saudáveis ainda pode ser questionável.
![]() No caso das dietas terapêuticas, destinadas à cães e gatos cardiopatas e hepatopatas, a sua inclusão possui maior respaldo científico.
No caso das dietas terapêuticas, destinadas à cães e gatos cardiopatas e hepatopatas, a sua inclusão possui maior respaldo científico.
Importância da inclusão de taurina e L-carnitina na dieta de cães e gatos
Caroline Fredrich Dourado Pinto1, Luciano Trevizan2
1Zootecnista e Doutoranda em Zootecnia, PPG Zootecnia – Universidade Federal do Rio Grande do Sul (UFRGS)
2Professor do Departamento de Zootecnia – UFRGS
  Leia também! Leia também!Insetos na alimentação pet: benefícios nutricionais e ambientais |
Blanchard, G.; Paragon, B. M.; Milliat, F.; Lutton, C. 2002. Dietary L-carnitine supplementation in obese cats alters carnitine metabolism and decreases ketosis during fasting and induced hepatic lipidosis. The Journal of Nutrition 132(2):204-210.
Center, S. A.; Harte, J.; Watrous, D.; Reynolds, A.; Watson, T. D.; Markwell, P. J.; Millington, D. S.; Wood, P. A.; Yeager, A. E.; Erb, H. N. 2000. The clinical and metabolic effects of rapid weight loss in obese pet cats and the influence of supplemental oral L-carnitine. Journal of Veterinary Internal Medicine 14(6):598–608.
Costa, N. D.; Labuc, R. H. Case report: efficacy of oral carnitine therapy for dilated cardiomyopathy in boxer dogs. 1994. The Journal of Nutrition 124(12 Suppl):2687S-2692S.
Epp, T. S.; Erickson, H. H.; Woodworth, J.; Poole, D. C. 2007. Effects of oral L-carnitine supplementation in racing greyhounds. Equine and Comparative Exercise Physiology 4:141–147.
Gross KL, Wedekind KJ, Kirk CA, et al. 1998. Effect of dietary carnitine or chromium on weight loss and body composition of obese dogs. Journal of Animal Science 76(Suppl. 1):175.
Hayes, K. C.; Carey, R. E.; Schmidt, S. Y. 1975. Retinal degeneration associated with taurine deficiency in the cat. Science. 188(4191):949-951.
Keene BW, Panciera, D. P.; Atkins, C. E.; Regitz, V.; Schmidt, M. J.; Shug, A. L. 1991. Myocardial L-carnitine deficiency in a family of dogs with dilated cardiomyopathy. Journal of the American Veterinary Medical Association, 198(4):647-650.
Kerner, J. & Hoppel, C. 2000. Fatty acid import into mitochondria. Biochimica et Biophysica Acta 1486:1-17.
McCusker, S.; Buff, P. R.; Yu, Z.; Fascetti, A. J. 2014. Amino acid content of selected plant, algae and insect species: a search for alternative protein sources for use in pet foods. Journal of Nutritional Science 3(e39):1-5.
Morris, J. G.; Rogers, Q. R.; Kim, S. W.; Backus, R. C. 1994. Dietary taurine requirement of cats is determined by microbial degradation of taurine in the gut. Taurine in Health and Disease. p.59-70. Springer, Boston, MA.
Pion, P. D.; Kittleson, M. D.; Rogers, Q. R.; Morris, J. G. 1987. Myocardial failure in cats associated with low plasma taurine: A reversible cardiomyopathy. Science. 237(4816):764-767.
Pion, P. D.; Kittleson, M. D.; Skiles, M. L.; Rogers, Q. R.; Morris, J. G. 1992a. Dilated cardiomyopathy associated with taurine deficiency in the domestic cat: relationship to diet and myocardial taurine content. Taurine. p.63-73. Springer, Boston, MA.
Pion, P. D.; Kittleson, M. D.; Thomas, W. P.; Skiles, M. L.; Rogers, Q. R. 1992b. Clinical findings in cats with dilated cardiomyopathy and relationship of findings to taurine deficiency. Journal of the American Veterinary Medical Association 201(2):267-274.
Pion, P. D.; Sanderson, S. L.; Kittleson, M. D. 1998. The effectiveness of taurine and levocarnitine in dogs with heart disease. Veterinary Clinics of North America: Small Animal Practice 28(6):1495-1514.
Rebouche, C. J.; Engel, A. G. 1983. Kinetic compartmental analysis of carnitine metabolism in the dog. Archives of Biochemistry and Biophysics 220:60-70.
Center, S. A.; Warner, K. L.; Randolph, J. F.; Sunvold, G. D.; Vickers, J. R. 2012. Influence of dietary supplementation with L-carnitine on metabolic rate, fatty acid oxidation, body condition and weight loss in overweight cats. American Journal of Veterinary Research 73(7):1002-1015.
Sanderson, S. L.; Osborne, C. A.; Lulich, J.P.; Gross, K. L.; Lowry, S. R.; Pierpont, M. E.; Ogburn, P. N.; Koehler, L. A.; Swanson, L. L.; Bird, K. A.; Ulrich, L. K. 2001. Effects of dietary fat and carnitine on urine carnitine excretion in dogs. Veterinary Therapeutics 2:181-192.
Spitze, A. R.; Wong, D. L.; Rogers, Q. R.; Fascetti, A. J. 2003. Taurine concentrations in animal feed ingredients; cooking influences taurine content. Journal of Animal Physiology and Animal Nutrition 87(7-8):251-262.
Sturman, J. A.; Gargano, A. D.; Messing, J. M.; Imaki, H. 1986. Feline maternal taurine deficiency: Effect on mother and offspring. The Journal of Nutrition 116(4):655-667.
Sturman, J. A.; Moretz, R. C.; French, J. H.; Wisniewski, H. M. 1985. Taurine deficiency in the developing cat: persistence of the cerebellar external granule cell layer. Journal of Neuroscience Research 13(3):405-416.
Varney, J. L.; Fowler, J. W.; Gilbert, W. C.; Coon, C. N. 2017. Utilisation of supplemented L-carnitine for fuel efficiency, as an antioxidant, and for muscle recovery in Labrador retrievers. Journal of Nutritional Science 6(e8):1-9.
Yue, K. T. N.; Fritz, I. B. 1962. Fate of tritium-labeled carnitine administered to dogs and cats. American Journal of Physiology 202:122-128.
[/registrados]










