

Generalmente se considera que la vacunación es el proceso más importante que resulta en la protección inmune de un lote. Esta percepción se deriva de la especificidad de este proceso.
Sin embargo, la comprensión actual de los principios inmunes implica que otros procesos de manejo sean muy relevantes en la mejora de la productividad a través del control de la inmunidad.
Inmunidad poblacional
- La inmunidad poblacional implica:
- Inversiones en calidad nutricional
- Uso racional de aditivos de rendimiento
- Uso de antibióticos
- Bioseguridad
- Capacitación de la fuerza de laboral
- Protocolo de vacunación bien estructurado
Estructura del MALT
El tejido linfoide asociado a la mucosa (MALT) está muy desarrollado. El componente intestinal de este sistema es el tejido linfoide asociado al intestino (GALT), que corresponde al 80% de todo el MALT y está compuesto por una compleja organización de órganos linfoides primarios y secundarios.
Las placas de Peyer (tejidos linfoides organizados presentes en la pared intestinal) están compuestos por linfocitos B, la mayoría de los cuales secretan IgA en el lumen intestinal.
Estas células componen 40% de las placas de Peyer:
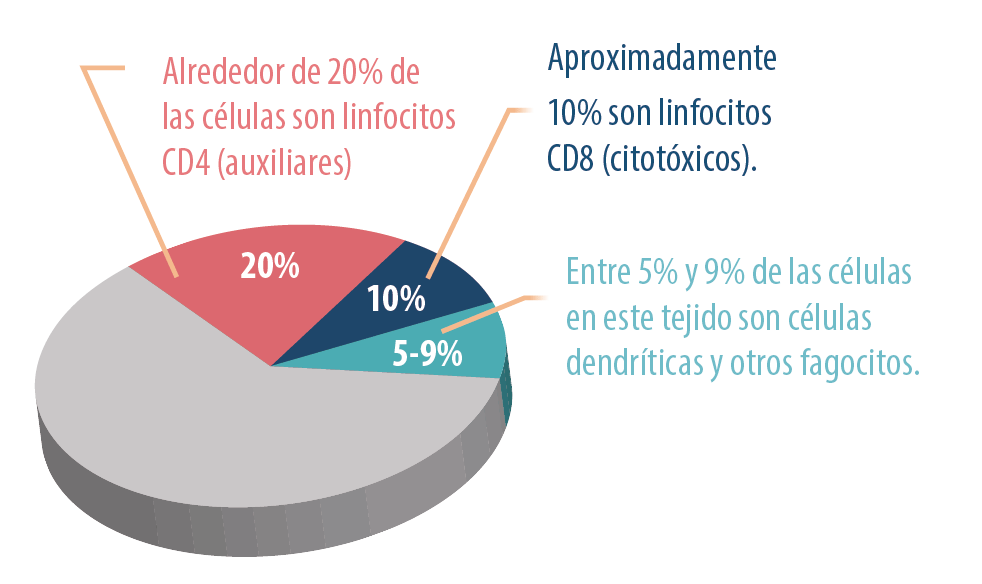

Las aves jóvenes presentan aproximadamente seis PPs, que al igual que otros componentes linfoides, están involucrados con la edad. En etapas posteriores de la vida, sólo se puede encontrar un agregado linfoide intestinal. Las amígdalas y los PP son fácilmente identificables en pollos de 10 días que alcanzan su máximo desarrollo entre las 5 y 16 semanas de edad. Con la involución natural relacionada con la edad, estos tejidos inmunes intestinales pueden no ser visibles a las 20 semanas.

Es un remanente del saco vitelino que muestra centros germinales con linfocitos B y macrófagos. La población de linfocitos intraepiteliales en el divertículo aumenta de 3 a 5 veces 5 días después de la incubación y hasta 10 veces a los 14 días y aún más hasta los 40 días de edad.
En las aves de corral, las células inmunes en los intestinos no se agrupan exclusivamente en los tejidos linfoides.
La mucosa intestinal, compuesta por el epitelio y la lámina propia, también es rica en leucocitos:
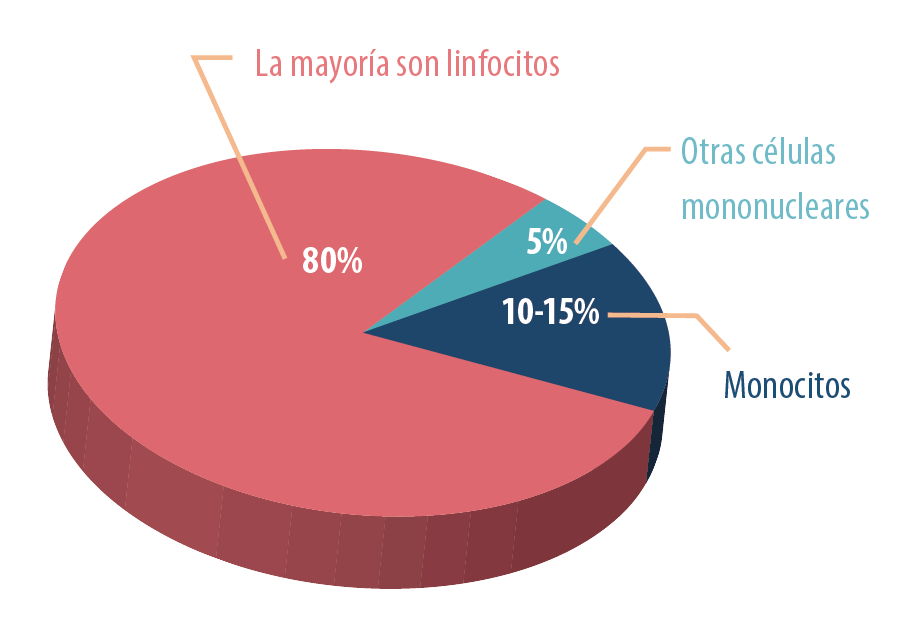

Estos linfocitos están presentes en el intestino en el momento de la eclosión, ya que lo han colonizado desde aproximadamente 16 días de la incubación. Entre los 4 y 6 días de vida esta colonización aumenta y alcanza la madurez en las primeras dos semanas de vida.
El procesamiento de materiales extraños (antígenos) por el GALT sigue una secuencia similar a la de los tejidos linfoides sistémicos. Sin embargo, en los enterocitos GALT también juegan un rol en el transporte de moléculas desde patógenos a los alcances de las células inmunes.
En el revestimiento epitelial intestinal, las células M son cruciales en la realización de esta tarea. Tanto los enterocitos como las células M contribuirán a las defensas de los patógenos.
La arquitectura intestinal local a menudo cambia en respuesta a patógenos, como alteraciones en la profundidad de las criptas, producción de moco, infiltración de células linfoides, aumento del espaciamiento entre las células, y así sucesivamente.
Se construirán respuestas más específicas en las placas de Peyer, donde los linfocitos B comenzarán las respuestas de IgA dirigidas contra el patógeno.
Inmunidad y nutrición
Después de la invasión de los patógenos, se producen muchos cambios estructurales en el intestino, relacionados con la permeabilidad, la infiltración celular, mejoramiento de las criptas, la producción de moco y enzimas, adicionado a las respuestas antigénicas específicas.[registrados]
Las respuestas específicas a los patógenos, denominada la respuesta inmune adaptativa, implican dos tipos principales de células:
Linfocitos B, que expresan inmunoglobulinas de superficie, y reaccionan con la producción de diferentes anticuerpos (respuesta inmune humoral).
Linfocitos T, que reconocen el complejo principal de histocompatibilidad (MHC) -complejo antígeno-MHC, presentado sobre la superficie de las células presentadoras de antígenos (macrófagos, células dendríticas y, finalmente, incluso enterocitos).
La captura de antígenos por pinocitosis o fagocitosis por las células M también determina la sensibilización de los linfocitos T y B en los centros germinales de PP. Después de la unión del antígeno a los receptores de células B, la división celular y la expansión clonal comienzan con la producción de anticuerpos específicos para el antígeno que incita a las células.
Debido a la conexión íntima de estos tejidos linfoides con el revestimiento epitelial intestinal, los componentes nutricionales tienen un gran efecto sobre las células inmunes locales. Por lo tanto, la integridad intestinal es tan relevante como los protocolos de vacunación para inducir una inmunidad adecuada a los desafíos ambientales. Entre más animales sean resistentes a las infecciones, menor será la transmisión lateral de patógenos seguido de un caso inicial dentro de la manada.


No es sorpresivo observar que la privación de alimentos afecta su maduración, aunque aparentemente obvia, ésta es una práctica bastante común en algunas unidades de producción. La ventana desde el nacimiento hasta el primer alimento que se observa frecuentemente en algunas empresas, asociada con las prácticas de vacunación en las plantas de incubación y el transporte hasta el alojamiento, puede generar una privación de alimento bastante extensa. En las primeras 48 a 72 horas de vida esto generará pérdidas importantes en el desarrollo del intestino.
La pérdida fisiológica que proviene del retraso en la alimentación se convierte en una pérdida económica directa e irrecuperable, reflejando no sólo un menor aumento de peso en los pollos de engorde, sino también un “costo inmune” para las aves incluso hasta las 60 semanas de edad o más.
Entre todos los factores que son relevantes en la protección natural de la manada, la composición óptima de la dieta determinará los mejores resultados posibles en el balance de los costos de activación de la inmunidad y la productividad animal.
La microbiota es obviamente muy relevante en este contexto. Los animales con un desafío ambiental extremadamente bajo (en condiciones de laboratorio) tienden a demostrar un rendimiento mejorado en comparación con las condiciones de campo, debido a los costos naturales de responder al desafío mediante la activación de la inmunidad.
Los animales libres de gérmenes tienen 10%-30% menos demanda metabólica que los porcinos convencionales. Sólo cuando la dieta es manejada de manera estricta y la microbiota es mejorada, los animales en condiciones normales pueden superar a los animales libres de gérmenes.
“Libre de gérmenes” no es una opción para los animales de granja. Es muy importante controlar las bacterias que interactúan con la inmunidad de la mucosa. Entre los dos puntos posibles para controlar la inmunidad intestinal (nutrición y microbiota), este último es mucho menos conocido.
Las respuestas a los “potenciadores de la microbiota” son usualmente inestables y varían ampliamente entre situaciones diferentes. Las mejores provienen del establecimiento de una microbiota de alta calidad desde las primeras semanas de vida, cuando las poblaciones bacterianas intestinales aún son lábiles. Como el impacto de la microbiota en la inmunidad y las interacciones dinámicas que crean son muy sensibles, actualmente se ha dedicado un capítulo de inmunología aviar a esto. El notable papel de los antibióticos ha sido y es (donde están permitidos ) un ejemplo obvio.
Los antibióticos promotores del crecimiento (potenciadores del rendimiento) tienen un tremendo impacto en la función inmune del intestino al alterar la constitución microbiana y, por lo tanto, funcionan como potenciadores del rendimiento a largo plazo.
Toda la cascada inmune se cambiará para tener el costo más bajo durante la vida del ave, si estos productos se usan para inducir una maduración óptima GALT.


La maduración inducida por antígeno del GALT puede seguir vías muy distintas, dependiendo del nivel y la calidad del estímulo que se proporciona.
En una situación equilibrada, la interacción antígeno-huésped conducirá a la producción de IgA intestinal. Esto se mantiene en un estado “saludable” por la acción de células supresoras y reguladoras que son abundantes en los tejidos de la mucosa.

En estos sitios, es común que las células dendríticas y linfoides secreten constantemente factores reguladores (como el retinol) que impiden las respuestas abiertas contra los antígenos orales, siendo crucial para evitar la inmunopatología: los tejidos de la mucosa no deben responder a la alimentación como lo harían a un patógeno, o de lo contrario surgiría alergia al alimento.
Por otro lado, los estímulos patógenos inducen fuertes respuestas inflamatorias intestinales, y los costos y el estrés involucrados en estas reacciones son escasos a la situación ideal. Por lo tanto, en este sentido, los antibióticos promotores de crecimiento tienen un gran impacto en la inmunidad y, por lo tanto, en la productividad.

Algunas moléculas inflamatorias pueden alcanzar el hipotálamo y controlar la temperatura corporal, induciendo fiebre. Esta es una respuesta antipatógena natural, pero está demandando mucha energía para aumentar la temperatura a unos pocos grados.
La inflamación también regula negativamente el apetito y el consumo de alimento disminuirá hasta que se alcancen las últimas etapas de inmunidad. Las fases posteriores de la inmunidad (como la producción de IgA), si bien son costosas, son menos exigentes en términos de “oportunidad”: no hay fiebre ni cambios en el apetito en estas fases.
Los “costos” generados por la inmunidad en el huésped son de dos naturalezas:
Las células inmunes tienen altas demandas de proteínas. No sólo eso, sino que la composición amino de las proteínas inmunes se desvía ampliamente de otras proteínas en el animal y, por consiguiente, las demandas nutricionales variarán si la inmunidad se activa constantemente. Como ejemplo, la valina y treonina se consumen en niveles más altos para la producción de IgA y de mucina (el constituyente principal del moco).
Las fases iniciales de las respuestas inmunes presentan un “costo de oportunidad” para el huésped. La protección inmunológica comienza con la inflamación.


En consecuencia, el objetivo al influir en la inmunidad de la mucosa es mantener los beneficios de la presencia de células inmunocompetentes y de IgA con una modulación mantenida de manera estricta, necesaria para regular los mediadores inflamatorios.
De esta manera, se puede explorar la respuesta inmune para actuar exitosamente contra cualquier encuentro futuro con los desafíos en el medio ambiente, al tiempo que se evita la pérdida de rendimiento relacionada con la inflamación.
En los últimos años, el aumento en el conocimiento de la dinámica involucrada en la respuesta inmune de la mucosa ha permitido el desarrollo de medidas que, especialmente para las aves de corral, pueden optimizar la producción, garantizando un equilibrio sensible entre la respuesta inmune y el rendimiento.
Por lo tanto, la nutrición es especialmente relevante para los animales en entornos desafiantes. Las respuestas a los patógenos en estas situaciones se producirán principalmente en la mucosa, en los intestinos y en el tracto respiratorio.
Controlar la inmunidad, ya sea a través de la microbiota o directamente por inmunonutrición, es vital para prevenir pérdidas de productividad. Se debe alcanzar un equilibrio entre las respuestas inmunitarias fuertes y el rendimiento. Esto depende de las interacciones involucradas en el órgano inmune más grande, el INTESTINO.

