



| Las pasturas templadas, representadas principalmente por gramíneas C3, leguminosas, y sus mezclas tienen cualidades innegables para ser utilizadas en la alimentación de rumiantes, incluso aquellos de leche o carne de alto potencial productivo. |


Por ejemplo, es conocido el aumento de ácidos grasos beneficiosos para la salud humana tanto en leche (Vibart et al., 2008; Morales-Almaraz et al., 2010; Mendoza et al., 2016a; Pastorini et al., 2019; Santana et al., 2023) como en carne (Cabrera y Saadoun, 2014; Fernández-Turren et al., 2020).
Por otra parte, cuando se consumen en su momento óptimo poseen características nutricionales de gran valor.


Algunas características de sus componentes químicos


|
Cuando esto último no sucede, la baja utilización de los compuestos proteicos en el rumen puede resultar en un desperdicio y un factor potencialmente contaminante (Ledgard et al., 1999; Totty et al., 2013). En sistemas de base pastoril, este desperdicio es difícil de evitar sin atentar contra la producción, ya que justamente cuando las pasturas tienen mejor calidad poseen más compuestos nitrogenados degradables.
|

En la literatura se reportan numerosos intentos para bajar la degradación ruminal de los compuestos nitrogenados de las pasturas utilizando herramientas como los taninos que tienen la cualidad de precipitar proteínas y transformarlas en proteína de pasaje (Orlandi et al., 2015; Pozo et al., 2022), a lo que se suma su potencial para reducir las emisiones de metano (Naumann et al., 2017), aunque tienen como desventaja la baja palatabilidad.
Una posible estrategia para aprovechar los compuestos nitrogenados de la pastura es el manejo de los horarios de pastoreo.
El ingreso en horas de la tarde, cuando las pasturas tienen más azúcares y menos humedad, tiene dos ventajas:
![]()

Aumentar el consumo de materia seca de la pastura, tanto por mejora en la palatabilidad dada por los azúcares,
![]()

como por un menor contenido de humedad (Huntington y Burns, 2007; Pozo et al. 2022).
La cantidad de azúcares y otros carbohidratos de rápida fermentación es variable para las distintas especies herbáceas y su acumulación depende de las horas luz, en general representa entre un 5 y un 10 % de la MS, concentración nada despreciable si los consideramos un “aditivo natural” para potenciar la fermentación ruminal.
Por otra parte, la degradabilidad ruminal de las fracciones fibrosas de este tipo de pastura es del orden del 50-60% (Cajarville et al., 2006a; Cajarville et al., 2012), valores altos considerando que se trata de las porciones menos degradables de los forrajes.




¿Y cómo es el ambiente ruminal de animales consumiendo pasturas de alta calidad?
Consistente con la elevada fermentación de los componentes químicos, el consumo de pasturas templadas de alta calidad resultará en concentraciones también elevadas de ácidos grasos volátiles (AGV) en el rumen.


El ambiente ruminal de animales consumiendo pasturas de buena calidad contrasta con el clásicamente reportado en la bibliografía para dietas forrajeras ricas en celulosa y pobres en almidones y azúcares,, con contenidos de proteína mucho más bajos, que resultan en bajas producciones de proteína microbiana y de AGV, con una elevada proporción de acetato dentro de los AGV (Church, 1993; Van Soest, 1994).
![]()

Las características de alta fermentación ruminal y por lo tanto, de alta producción de AGV, promueven, cuando la pastura no es limitante en cuanto a cantidad, ambientes ruminales que se caracterizan por valores de pH relativamente bajos (Figura 1), fundamentalmente en los horarios posteriores a pastoreos intensos.
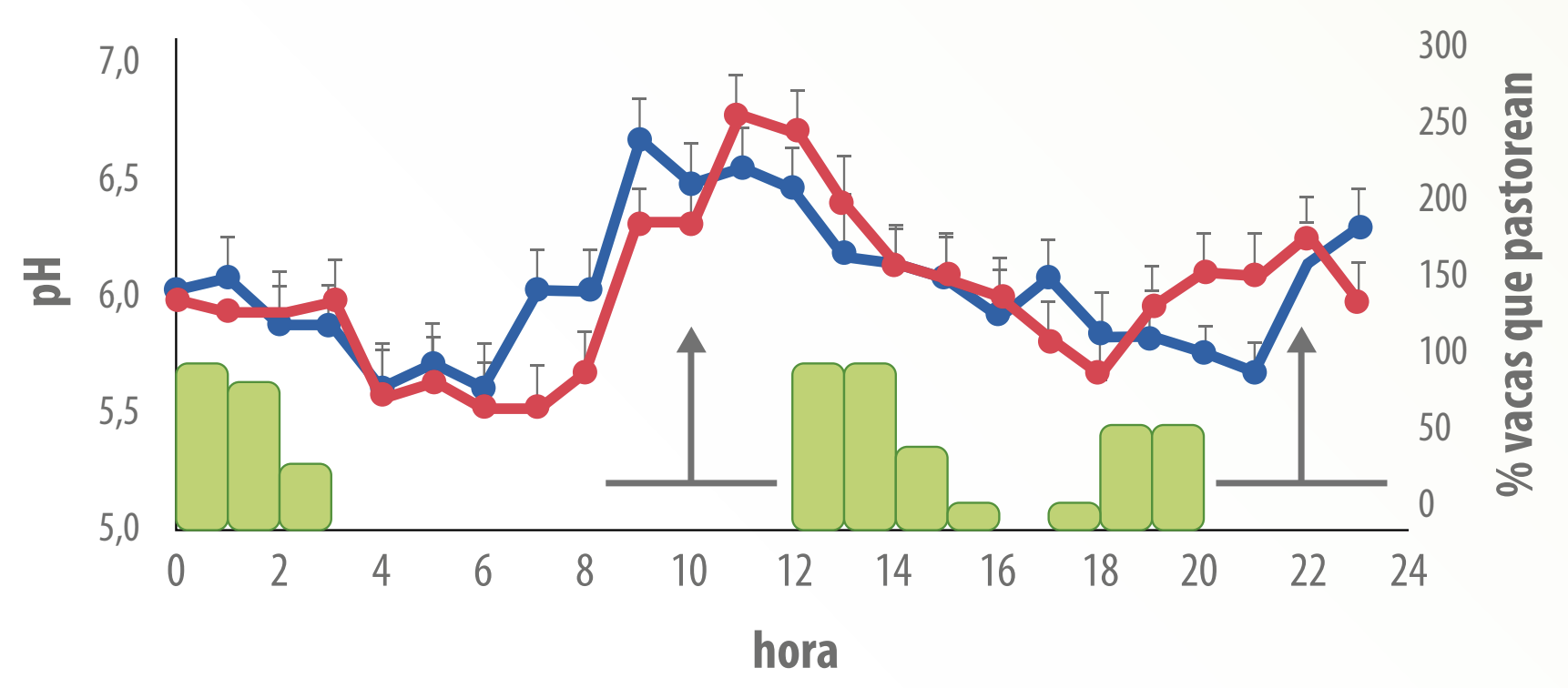

Figura 1. Evolución del pH ruminal a lo largo del día en vacas en pastoreo suplementadas con grano de trigo (curva azul) o grano de maíz (curva roja). Las columnas verdes representan la proporción de vacas pastoreando luego del período de confinamiento. Las flechas indican el momento de la suplementación y las líneas bajo las flechas indican los períodos de confinamiento. Es interesante observar que el pH ruminal de los animales baja cuando inician los horarios de pastoreo intenso, y no luego de la suplementación con granos (Extraído de Cajarville et al., 2006a).


|
En ocasiones, estos pH llegan a valores que pueden considerarse, según los autores (Brossard et al., 2003; Krauze y Oetzel, 2006) propios de acidosis subclínica (SARA). En un análisis de datos provenientes de varios experimentos con animales consumiendo dietas con predominio de pasturas templadas mixtas (gramíneas y leguminosas) suministradas frescas (Figura 2), observamos que el tiempo de acceso al alimento tiene una influencia importante en el pH ruminal y su dinámica. |
Los animales con acceso continuo al alimento presentaron valores de pH más altos y estables a lo largo del día independientemente si habían recibido o no suplementación con concentrados. Por el contrario, cuando el acceso a los alimentos era restringido, con largas horas de ayuno o suministrado en pocas comidas diarias, los pH ruminales transcurrían en valores inferiores a 6 durante muchas horas al día, por lo que el riesgo de entrar en una condición de SARA es elevado.
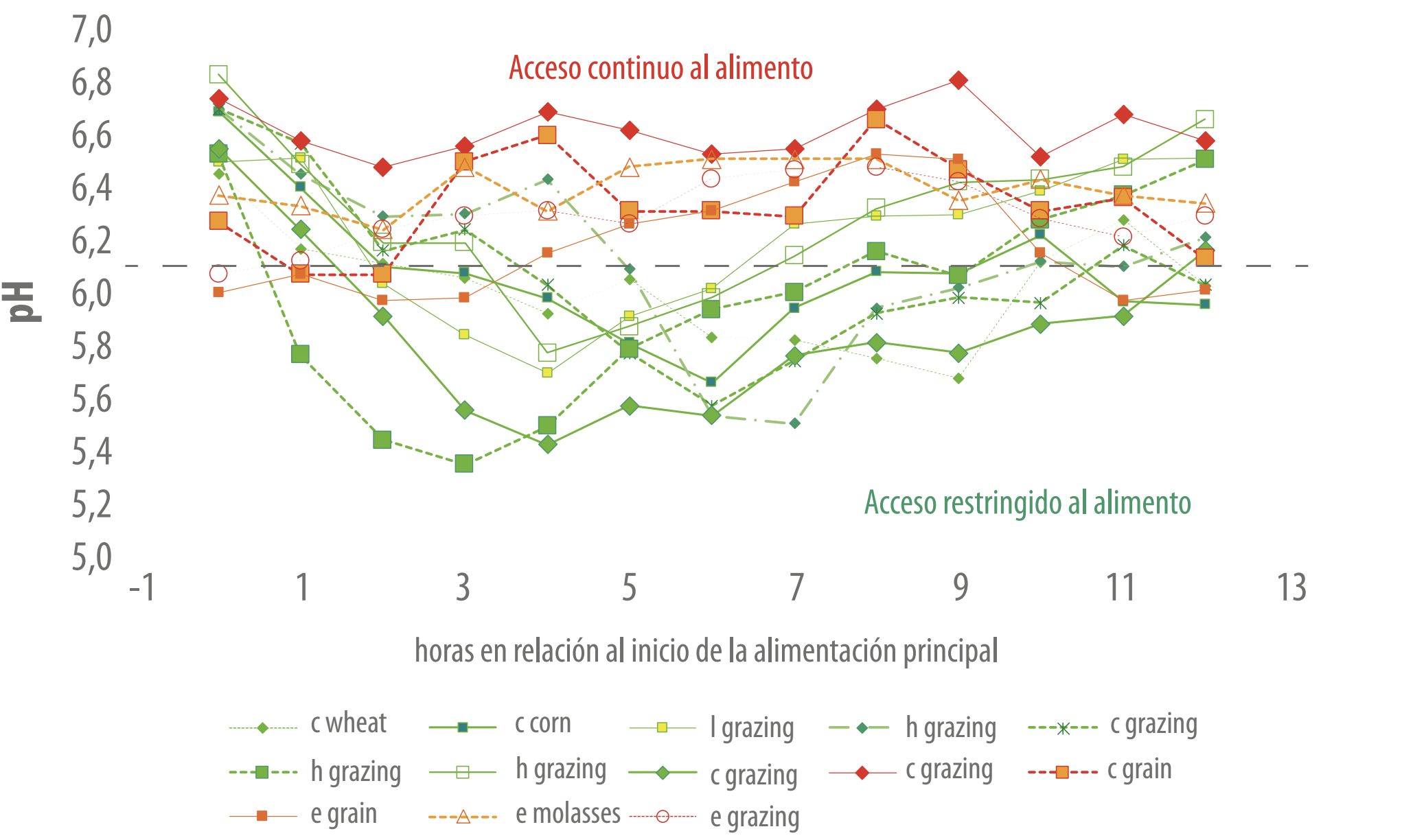

Figura 2. Dinámica de pH ruminal de animales correspondientes a 1392 mediciones de pH realizadas en un total de 58 animales, distribuidos en 13 tratamientos experimentales con dietas a base de pasturas templadas suministradas frescas. Las curvas de color verde indican animales que tenían acceso restringido al alimento, las rojas indican animales que tenían acceso continuo al alimento. La línea blanca indica el pH que se considera umbral para asegurar el mantenimiento de la salud del rumen (6-6,2). Observar cómo se separan las curvas verdes de las rojas justamente en torno al pH umbral, independiente del tipo de animal y del tipo de alimentación (Códigos empleados: c=vacas, h=terneras de recría/vaquillonas, l=corderos, e=ovejas adultas; grazing=sólo pastoreo, o suplementación a razón de 1/3 de la dieta total con: corn=maíz, wheat=trigo, molasses=melaza de caña de azúcar, grain=mezcla de granos. Datos extraídos de Cajarville et al. (2006c)).
Félix et al. (2017) en un experimento posterior con vaquillonas en crecimiento consumiendo sólo pasturas de alta calidad con predominio de gramíneas, constatan la inestabilidad ruminal y el ingreso menor de energía al animal a medida que las horas de ayuno se alargan.
Es de hacer notar, que las leguminosas podrían tener un efecto “preventivo” sobre las bajadas excesivas de pH que se producen cuando la ingesta de pastura es especialmente intensa (Pérez-Ruchel et al., 2013), por ejemplo, cuando las vacas lecheras salen a pastorear luego del ordeñe, o cuando abrimos una nueva franja en pastoreos intensivos. Lo anterior se explica por la alta capacidad tampón de las leguminosas, cuyos efectos son positivos a nivel ruminal.
Las pasturas templadas como fuente de la mejor proteína para alimentar al rumiante
Recordemos cómo es la alimentación proteica de los rumiantes. Gran parte de los microorganismos sintetizados en el rumen pasan al tracto digestivo posterior, donde se digieren y absorben, constituyendo la mayor proporción (más del 70%) de la proteína metabolizable que se absorbe y alimenta al rumiante.


| La alta fermentescibilidad y el alto contenido en proteínas degradables de las pasturas de buena calidad, promueven un activo crecimiento y desarrollo de la microbiota a nivel ruminal.
Por ello, indirectamente (a través de la proteína microbiana formada), resultan en una excelente fuente de proteína de alta calidad para los rumiantes. |
La alta fermentescibilidad y el alto contenido en proteínas degradables de las pasturas de buena calidad, promueven un activo crecimiento y desarrollo de la microbiota a nivel ruminal. Por ello, indirectamente (a través de la proteína microbiana formada), resultan en una excelente fuente de proteína de alta calidad para los rumiantes.
Trabajos realizados en Argentina, Uruguay y el sur de Brasil, donde se emplean pasturas templadas de alta calidad en los sistemas pastoriles intensivos, muestran que la cantidad de proteína microbiana sintetizada en el rumen es alta en estas condiciones (García et al., 2000; Amaral et al., 2011; Tebot et al., 2012; Aguerre et al., 2013).
![]()

Esto quiere decir que, desde el punto de vista del uso del rumen como sistema de fermentación, el consumo de pasturas de buena calidad nos llevaría a un máximo nivel, aportando al rumiante una fuente proteica de calidad en importante cantidad.
De todas formas, la principal restricción para lograr una alta inclusión de pasturas en dietas de rumiantes de alta producción es debida a la limitación del consumo. Además de la dificultad que representan las actividades de búsqueda y caminata asociadas a la recolección de la pastura, que han sido responsabilizadas del bajo consumo en dietas pastoriles, algunas observaciones indican que las vacas comen con más lentitud la pastura que la ración totalmente mezclada, aun cuando la primera esté provista en comederos (Pomiés et al., 2015; Mendoza et al., 2018).




Para terminar…
En todo este artículo nos hemos referido a pasturas de alta calidad, pero nuestros lectores se preguntarán cómo hacemos para que nuestros animales consuman siempre este tipo de pasturas, manteniendo la calidad de una pastura ya instalada, aprovechando así su potencial.
|
Todos sabemos que la calidad de la pastura no depende solamente de las especies que la componen. Es muy habitual ver en establecimientos de base pastoril lecheros o ganaderos, sobre todo en épocas del año en que el crecimiento del pasto es más acelerado que la capacidad de consumo por parte de los animales, que los animales consumen materiales en estados de maduración avanzadas, comprometiendo la calidad de lo ingerido. Ello incluso se da en predios que cuentan con una base pastoril con “potencial” buena calidad. Para evitar este problema es fundamental tener sistemas de manejo del pastoreo preestablecidos que nos permitan adelantarnos al ritmo de crecimiento de la pastura. De esa forma lograremos un consumo de forraje de alta calidad, estable a lo largo del año. En esta misma edición presentamos un artículo referido al tema de manejo de pasturas, complemento indispensable para llevar a la práctica los conceptos que comentamos en este artículo (ver El cálculo del miedo) |
Bibliografía
Aguerre M., Cajarville C., Kozloski G. V, Repetto J.L. 2013. Intake and digestive responses by ruminants fed fresh temperate pasture supplemented with increased levels of sorghum grain: A comparison between cattle and sheep. Anim. Feed Sci. Technol. 186: 12–19.
Amaral, G. A., Kozloski, G. V., Santos, A. B., Castagnino, D. S., Fluck, A. C., Farenzena, R., Alves, T. P., & Mesquita, F. R. 2011. Metabolizable protein and energy supply in lambs fed annual ryegrass (Lolium multiflorum Lam.) supplemented with sources of protein and energy. J. Agric. Sci. 149, 4: 519–527.
Bargo F., Muller L.D., Varga G.A., Delahoy J.E., Cassidy T.W. 2002. Ruminal digestion and fermentation of high-producing dairy cows with three different feeding systems combining pasture and total mixed rations. J. Dairy Sci. 85: 2964–2973.
Berthiaume R., Tremblay G., Castonguay Y., Bertrand A., Bélanger G., Lafrenière C., Michaud R. 2006. Length of the daylight period before cutting improves rumen fermentation of alfalfa assessed by in vitro gas production. J. Anim. Sci. 84 (Suppl .1): 102.
Brossard L., Martin C., Michalet-Doreau B. 2003. Ruminal fermentative parameters and blood acido-basic balance changes during the onset and recovery of induced latent acidosis in sheep. Anim. Res. 52: 513–530.
Cabrera M.C., Saadoun A. 2014. An overview of the nutritional value of beef and lamb meat from South America. Meat Sci. 98: 435–444.
Cajarville C., Aguerre M., Repetto J.L. 2006a. Rumen pH, NH3-N concentration and forage degradation kinetics of cows grazing temperate pastures and supplemented with different sources of grain Anim. Res. 55: 511-520.
Cajarville C., Pérez A., Aguerre M., Britos A., Repetto J.L. 2006b. Effect of the timing of cut on ruminal environment of lambs consuming temperate pastures. J. Dairy Sci. 89, (Suppl. 1): 103.
Cajarville C., Aguerre M., Britos A., Tebot I., Pérez A., Elizondo V., Repetto J.L. 2006c. Effect of feeding frequency of fresh forage on ruminal pH: data review. Proc. of the 14th International Symposium on Lameness in Ruminants. 242 pp.
Cajarville C., Britos A., Garciarena D., Repetto J.L. 2012. Temperate forages ensiled with molasses or fresh cheese whey: Effects on conservation quality, effluent losses and ruminal degradation. Anim. Feed Sci. Technol. 171: 14-19.
Cajarville C., Britos A., Errandonea N., Gutierrez L., Cozzolino D., Repetto J.L. 2015. Diurnal changes in water-soluble carbohydrate concentration in Lucerneand tall fescue in autumn and the effects on in vitro fermentation. New Zeal. J. Agric. Res. 58: 281-291.
Church D.C. 1993. El rumiante: fisiología digestiva y nutrición. Zaragoza, Ed. Acribia, 641pp.
Cohen D.C. 2001. Degradability of crude protein from clover herbages used in irrigated dairy production systems in Northern Victoria. Aust. J. Agric. Res. 52: 415-425.
Felix A., Repetto J.L., Hernández N., Pérez-Ruchel A., Cajarville C. 2017.Restricting the time of access to fresh forage reduces intake and energy balance but does not affect the digestive utilization of nutrients in beef heifers. Anim. Feed Sci. Technol. 226: 103-112.
Fernandez-Turren G., Repetto J.L., Arroyo J.M., Pérez-Ruchel A., Cajarville C. 2020. Lamb Fattening Under Intensive Pasture-Based Systems: A Review. Animals 10: 382.
García S.C., Santini F.J., Elizalde J.C. 2000. Sites of digestion and bacterial protein synthesis in dairy heifers fed fresh oats with or without corn or barley grain. J. Dairy Sci. 83: 746-755.
Huntington G.B., Burns J.C. 2007. Afternoon harvest increase readily fermentable carbohydrate concentration and voluntary intake of gamagrass and switchgrass baleage by beef steers. J. Anim. Sci. 85: 276-284
Kokko C., Soder K.J., Brito A.F., Hovey R.S., Berthiaume R. 2013. Effect of time of cutting and maceration on nutrient flow, microbial protein synthesis, and digestibility in dual-flow continuous culture. J. Anim. Sci. 91:1765-1774.
Krause K.M., Oetzel G.R. 2006. Understanding and preventing subacute ruminal acidosis in dairy herds: a review. Anim. Feed Sci. Technol. 126: 215-236.
Ledgard S.F., Penno J.W., Sprosen M.S. 1999. Nitrogen inputs and losses from clover/grass pastures grazed by dairy cows, as affected by nitrogen fertilizer application. J. Agric. Sci.Camb. 132: 215–225
Mee J.F., Boyle L.A. 2020. Assessing whether dairy cow welfare is “better” in pasture-based than in confinement-based management systems, New Zeal. Vet. J. 68: 168-177.
Mendoza A., Cajarville C., Repetto J.L. 2016a. Short communication: Intake, milk production, and milk fatty acid profile of dairy cows fed diets combining fresh forage with a total mixed ration. J. Dairy Sci. 99:1938–1944
Mendoza A., Cajarville C., Repetto J.L. 2016b. Digestive response of dairy cows fed diets combining fresh forage with a total mixed ration. J. Dairy Sci. 99:8779–8789.
Mendoza A., Cajarville C., Repetto J.L. 2018. Behaviour of cows fed a total mixed ration with different access time to fresh forage. New Zeal. J. Agric. Res. 61,1: 102-108.
Morales-Almaraz E., Soldado A., Gonzalez A., Martínez A., Domínguez I., de la Roza B., Vicente F. 2010. Improving the fatty acid profile of dairy cow milk by combining grazing with feeding of total mixed ration. J Dairy Res. 77: 225–230
Nápoli G.M., Santini F.J. 1988. Suplementación energético-proteica de forrajes frescos en condiciones de pastoreo: I. Efecto sobre el medio ambiente ruminal y la degradabilidad proteica. Rev. Arg. Prod. Anim. Vol. 8 (Sup.1): 39
Naumann H.D., Tedeschi L.O., Zeller W.E., Huntley N.F. 2017. The role of condensed tannins in ruminant animal production: advances, limitations and future directions. Rev. Bras. Zootec., 46: 929-949.
Oba M., Allen M.S. 1999. Evaluation of the Importance of the digestibility of neutral detergent fiber from forage: effects on dry matter intake and milk yield of dairy cows J. Dairy Sci. 82: 589–596.
Orlandi T., Kozloski G.V., Alves T.P., Mesquita F.R., Ávila S.C. 2015. Digestibility, ruminal fermentation and duodenal flux of amino acids in steers fed grass forage plus concentrate containing increasing levels of Acacia mearnsii tannin extract. Anim. Feed Sci. Technol. 210: 37-45.
Pastorini M., Pomiés N., Repetto J.L., Mendoza A., Cajarville C. 2019. Productive performance and digestive response of dairy cows fed different diets combining a total mixed ration and fresh forage. J. Dairy Sci. 102: 4118–4130.
Pérez-Ruchel A., Repetto J.L., Cajarville C. 2013. Suitability of live yeast addition to alleviate the adverse effects due to the restriction of the time of access to feed in sheep fed only pasture. J. Anim. Phys. Anim. Nutr. 97: 1043–1050.
Pomiés N., Pastorini M., Cajarville C., Mendoza A., Repetto J.L. 2015 Combinación de ración totalmente mezclada y forraje fresco efecto sobre el comportamiento ingestivo y la tasa de consumo de vacas lecheras. XXIV Reunión de la Asociación Latinoamericana de Producción Animal, Puerto Varas, Chile
Pozo C.A., Kozloski G.V., Cuffia M., Repetto J.L., Cajarville C. 2022. Changing the grazing session from morning to afternoon or including tannins in the diet was effective in decreasing the urinary nitrogen of dairy cows fed a total mixed ration and herbage. J. Dairy Sci.105: 4987-5003.
Repetto J.L., Cajarville C., D’Alessandro J., Curbelo A., Soto C., Garín D. 2005. Effect of wilting and ensiling on ruminal degradability of temperate grass and legume mixtures. Anim. Res. 54: 73-80
Santana A., Cajarville C., Mendoza A., Repetto J.L. 2023. Including 8 hours of access to alfalfa in 1 or 2 grazing sessions in dairy cows fed a partial mixed ration: Effects on intake, behavior, digestion, and milk production and composition. J. Dairy Sci. 106: 6060–6079.
Sauvant D., Grenet E., M-Doreau B. 1995. Dégradation chimique des aliments dans le réticulo-rumen: cinétique et importance. En: Nutrition des ruminants domestiques. Ed. INRA, Paris.
Tebot I., Cajarville C., Repetto J.L., Cirio A. 2012. Supplementation with non-fibrous carbohydrates reduced fiber digestibility and did not improve microbial protein synthesis in sheep fed fresh forage of two nutritive values. Animal 6, 4: 617–623.
Totty V.K., Greenwood S.L., Bryant R.H., Edwards G.R. 2013. Nitrogen partitioning and milk production of dairy cows grazing simple and diverse pastures. J. Dairy Sci. 96: 141–149.
Van Soest P.J. 1994. Nutritional ecology of the ruminant. 2a. ed., Ithaca, Ed. Cornell University Press, 476p.
Vibart R.E., Fellner V., Burns J.C., Huntington J.B., Green J.T. 2008. Performance of lactating dairy cows fed varying levels of total mixed ration and pasture. J. Dairy Res. 75: 471–480.
Vibart R.E., Burns J.C., Fellner V. 2010. Effect of replacing total mixed ration with pasture on ruminal fermentation. Prof. Anim. Sci. 26: 435–442.