



La metionina es el primer aminoácido limitante en aves de corral y uno de los primeros en otras especies productivas.
Las dietas a base de maíz y soja son deficitarias en este aminoácido, por lo tanto, la suplementación con metionina en alimentación animal se ha venido realizando de forma rutinaria desde hace tiempo.
Las primeras fuentes de metionina comercialmente disponibles:
- DL-Metionina
- Su hidroxianálogo (DL-HMTBA o MHA-FA o MHA-Ca).
Ambas se producen a partir de recursos no renovables mediante síntesis química, consecuentemente ninguna es una fuente natural de metionina. Debido a esto, tanto la D-Metionina como el DL-HMTBA necesitan ser transformadas a L-Metionina por el animal suponiendo un coste de energía, aminoácidos (para la aminación de la Keto-Metionina), actividad enzimática y celular (Figura 1).

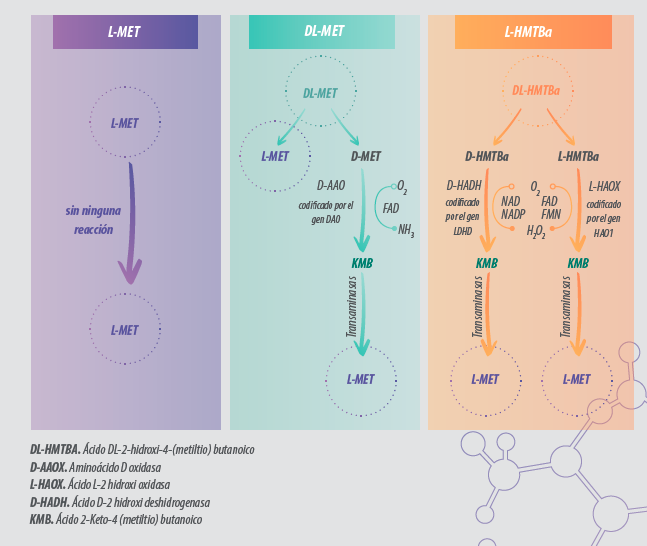
Figura 1. Metabolismo de las diferentes fuentes de metionina (Met) en la dieta. El isómero D-Met y el precursor Ácido DL-2-hidroxi-4-(metiltio) butanoico (DL-HMTBA) deben ser transformados en L-Met para su utilización. Diferentes enzimas y cofactores están implicados en este proceso (Zhang et al., 2018).
Sin embargo, actualmente tenemos disponible en el mercado volúmenes importantes de L-Metionina obtenida de recursos renovables y mediante fermentación.
Esta L-Metionina cristalina brinda la oportunidad de liberar a los animales del trabajo adicional e innecesario de convertir estos D-isómeros y DL-precursores en L-Metionina.


Los nutricionistas cuando utilizan estas fuentes de metionina siempre se enfrentan a la cuestión de su biodisponibilidad, y ésta puede variar dependiendo de cuál sea el interés, comercial o nutricional.
El profesor Baker es el principal científico citado cuando se habla sobre la biodisponibilidad de las fuentes de metionina (Katz and Baker, 1975). El propio Baker escribió que “la L-Metionina es mejor fuente de aminoácidos sulfurados que la D-Metionina”, reportando una biodisponibilidad del 90% para la D-Metionina en pollos (Baker, 1994).
En publicaciones más recientes, tanto Shen et al. (2015) como Esteve- García y Khan (2018), han descrito una biodisponibilidad más baja para la DL-Metionina comparada con la L-Metionina (71 y 77% vs 100% L-Met, respectivamente)en broilers modernos (Figura 2).

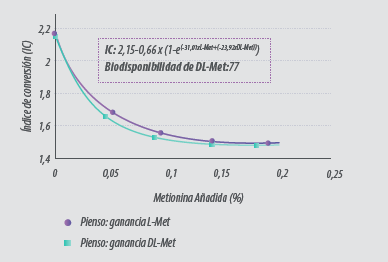
Figura 2. Índice de conversión (FCR) de pollos a los 37 días de edad alimentados con diferentes niveles de DL y L-Metionina.
Cuando hablamos del hidroxianálogo, según Sauer et al. (2008) el DL-HMTBA tiene una biodisponibilidad menor respecto a la DL-Met que varía entre el 45 y el 88% para el MHA-FA (12% de agua), dependiendo de:
- El parámetro estudiado (peso corporal, índice de conversión o rendimiento muscular de la pechuga).
- El nivel inicial de aminoácidos sulfurados.
- El modelo de estimación aplicado.
De media, comercialmente se aceptan niveles de biodisponibilidad de un 77% para el DL-HMBTA respecto a la DL-Metionina.
Las publicaciones realizadas por Saunderson (1985 y 1987) resultan realmente útiles para entender mejor el porqué de las diferencias entre las fuentes de metionina.


En un artículo científico publicado por Saunderson en 1985, se explicaban los resultados obtenidos tras administrar diferentes fuentes de metionina radiomarcadas en broilers. La excreción de las diferentes fuentes de metionina fue (Fig. 3):
- DL-Met excretada: un 10% del producto administrado.
- DL-HMTBA excretada: un 21% del producto administrado.
- L-Met: solo un 2% fue excretada.

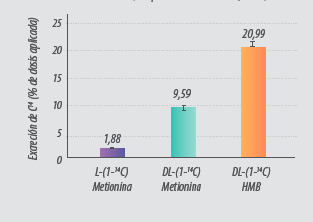
Figura 3. Excreción de L-Met, DL-Met y DL-HMB marcadas con 14C (adaptado de Saunderson, 1985).
Además la tasa de oxidación varió entre productos (Fig. 4):
- La DL-Met presentó la tasa de oxidación más alta (5,5% de la dosis dada).
- El DL-HMTBA se oxidó entre un 3 y un 7% (5.0 ± 1.8%)
- La oxidación de L-Met se limitó a un 4.2% de la dosis administrada.

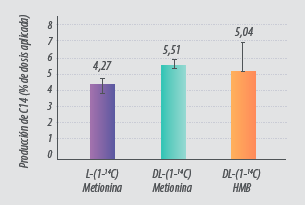
Figura 4. Oxidación de L-Met, DL-Met y DL-HMB marcadas con 14C (adaptado de Saunderson, 1985).
Curiosamente, la suma de la excreción, la oxidación y el contenido en agua del DL-HMTBA reduciría su biodisponibilidad a un 62% (Formula 1).


En el caso de la DL-Metionina, la suma de la excreción y la oxidación reducirían su biodisponibilidad a un 85% (Formula 2).


En el caso de la L-Metionina, ésta sigue las vías metabólicas naturales y estándar en los animales. De este modo, la L-Met también se excretó (1.8%) y se oxidó (4.2%) tras su administración.
Por consiguiente, debemos tener en consideración el metabolismo de la L-Metionina en un 6% (excreción y oxidación).
Siendo conservadores, esto nos lleva a unos valores de biodisponibilidad del 90.5 % para la DL-Metionina y del 67% para el DL-HMTBA respecto a la L-Metionina, sin tener en cuenta la eficacia de la metionina en la síntesis de proteína muscular y de otros órganos. De hecho, se observó una mayor deposición de L-Met en la pechuga y en los muslos en comparación con DL-Met y DL-HMTBA (Saunderson, 1985 y 1987)(Figura 5).

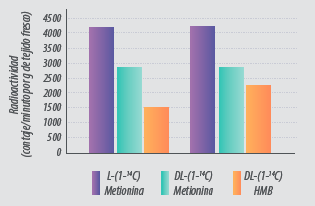
Figura 5. Deposición de L-Met, DL-Met y DL-HMB marcadas con 14C en proteína muscular (adaptado de Saunderson, 1985).

- Para valorar el posible efecto del nivel de alimentación o de los niveles de aminoácidos azufrados sobre la excreción de las diferentes fuentes de metionina, se diseñó un segundo estudio en el que los animales se sometieron a una deficiencia de metionina y una restricción de alimento (Saunderson, 1987), estudiándose los mismos parámetros.
- Los resultados obtenidos demostraron que la restricción alimentaria no varía los niveles de excreción de las diferentes fuentes de metionina.
- Por otro lado, la oxidación de la DL-Metionina sí que aumentó hasta el 10% de la dosis administrada en condiciones de alimentación restringida, siendo la tasa de oxidación más alta de las fuentes de metionina utilizadas.
En conclusión, el DL-HMTBA y la DL-Metionina podrían ser fácilmente reemplazables por un 67 % y 90.5 % de L-Metionina sin comprometer los parámetros productivos de los animales.