



El conocimiento de la microbiota que habita el sistema digestivo de las aves y su función metabólica es de gran importancia para mejorar su desempeño. La cantidad y la composición de la microflora varían en función de la edad del ave y del segmento intestinal en cuestión.
Colonización bacteriana del intestino
El óvulo proporciona al embrión todos los nutrientes y condiciones necesarias para su desarrollo a pollito, además del aislamiento del ambiente externo a través de la cáscara y la cutícula protectora que lo cubre. Sin embargo, se sabe que la cutícula sufre degradación con el tiempo y, a menudo, no se distribuye uniformemente sobre la cáscara del huevo, lo que podría facilitar el transporte de bacterias del entorno externo al interno.
Es muy probable que la puerta de entrada para los microorganismos dentro del huevo sean los poros, principalmente aquellos sin la cobertura adecuada de material proteico, los poros permeables. Después de la penetración en los poros, las bacterias pueden alcanzar otros canales (especialmente en la capa mamilar), donde hay humedad proveniente del huevo, lo que permite que se multiplique (Figura 1).
Figura 1. Estructuras del huevo
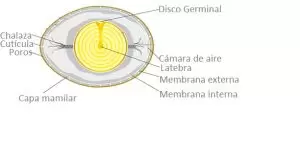

Los microorganismos que pasan a través de la cáscara alcanzan las membranas de la cáscara (externa e interna). La membrana interna es más eficiente para controlar la penetración bacteriana, pero los espacios que deja la conexión irregular entre las dos membranas permiten que las bacterias permanezcan y se reproduzcan.
Está ampliamente aceptado que los patógenos como Campylobacter y Salmonella experimentan una transmisión vertical desde la matriz al pollito. En este sentido, debe considerarse la colonización del tracto reproductivo de las gallinas, lo que significaría que el huevo fértil podría colonizarse antes de la puesta (Figura 2).
Al igual que el tracto reproductivo de la hembra, el macho también puede albergar bacterias. Por lo tanto sería justo considerar la hipótesis de que los microorganismos beneficiosos también podrían transmitirse verticalmente. Independientemente de la fuente primaria de contaminación, está confirmado que los intestinos de los pollitos comienzan a colonizarse incluso antes del nacimiento. [registrados]
Pedroso et al. (2008) encontraron la existencia de microbiota con baja diversidad en el intestino de embriones de pollos luego del día 16 de incubación. Las muestras tomadas en el momento de llegada de los pollitos a la granja también mostraron que el tracto intestinal de los mismos ya estaba completamente colonizado.
Luego de la eclosión, el pollito tiene contacto con los microorganismos de manera constante. Continuamente, la exposición bacteriana a través de la ración, el agua, la cama, el polvo y el personal, van a propiciar el desarrollo y la maduración de las comunidades microbianas.
Figura 2. Fuentes de contaminación con microorganismos durante el proceso de crianza de pollos.
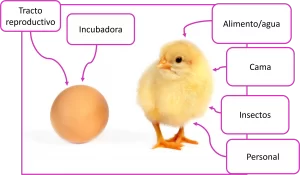

Adaptado de Macari et al., 2014
Inicialmente, la microbiota presente en el tracto gastrointestinal es inmadura y presenta una baja diversidad; hay una gran diferencia entre la composición bacteriana desde los 3 a los 49 días de edad, pero poca alteración entre 14, 21 e 28 días de edad (Lu et al., 2003). La microbiota presente en el intestino delgado de los pollitos progresa a los ciegos donde se establece y, después de la primera semana de vida, se diferencia, adquiriendo características propias. En la primer semana, las principales especies bacterianas observadas son Clostridium, Ruminococcus, Lactobacillus y Proteobacterias, que incluye las principales cepas patogénicas Gram-negativas, como Salmonella y Escherichia coli.
Microbiota residente y microbiota transitoria
- La microbiota residente también llamada microbiota normal está compuesta por bacterias, hongos y protozoarios. La microbiota normal por definición, se establece permanentemente en el huésped y no causa enfermedades en los individuos saludables.
- La microbiota transitoria puede ser definida como aquella que perdura en el tracto gastrointestinal por un período de tiempo y después desaparece.
Tanto la microbiota residente como la transitoria, pueden contener patógenos potenciales causadores de enfermedades si se dieran las condiciones propicias.
Macrohabitats
Buche, proventrículo y molleja
Las condiciones de pH ácido en el buche y, principalmente, en los estómagos glandular (proventrículo) y muscular (molleja) son la primera barrera natural para la entrada y la proliferación de bacterias patógenas, que normalmente están más adaptadas a condiciones de pH más neutro. Por este motivo, todo el tracto superior es colonizado exclusivamente por Lactobacillus y otras bacterias ácido-lácticas (Figura 3).


En el buche, hay una pequeña fermentación bacteriana, que lleva a cabo la hidrólisis de los carbohidratos, principalmente del almidón. En el proventrículo ocurre el inicio de la hidrólisis mediante la secreción de ácidos y enzimas, pero el tiempo de permanencia en esta porción es corto. En el estomago muscular ocurre la “molienda” de las partículas alimenticias y la mezcla con las secreciones estomacales, proporcionando mayor acción de las mismas. En esta porción, es posible una mayor acción de las enzimas microbianas, aumentando la disponibilidad de nutrientes para el ave. Además, las bacterias muertas en esta primera etapa, sufren la acción de los jugos gástricos y representan una pequeña fuente de proteína bacteriana para el animal.
Figura 3. Bacterias ácido-lácticas

Intestino delgado
Hay una distribución lógica de las comunidades bacterianas dentro del intestino delgado a lo largo de sus diferentes segmentos, debido a las condiciones ambientales intrínsecas de cada porción (nivel de nutrientes, pH, presencia de secreciones digestivas, etc.). El duodeno, a pesar de parecer inhóspito para el crecimiento bacteriano por la alta concentración salina y la gran variación de pH, presenta una cantidad de mucus extremamente espesa y, dentro de ésta, permite la colonización bacteriana, aunque la concentración es baja.
El yeyuno es la porción del intestino donde ocurre la mayor parte de la digestión propiamente dicha. La presencia de algunas cepas fermentadoras de carbohidratos estructurales, como Ruminococcus, ya comienzan a ser detectadas en este ambiente.
El íleon, es la porción final del intestino delgado. Las condiciones de pH aquí son más neutras (6,3-7,2) y la cantidad de mucus es más constante. Estos factores generan un número total de poblaciones bacterianas mayor. Las comunidades son dominadas por bacterias ácido-lácticas, principalmente Lactobacillus, pudiendo presentarse enterobacterias y Clostridium en mayor o menor cantidad, dependiendo de las condiciones dieteticas del animal.
Figura 4. Esporas de Clostridium

Ciegos
Los ciegos son estructuras pares, con forma sacular, dentro de las cuales el ambiente es de anaerobiosis y el alimento permanece un tiempo considerable. Estas características hacen que los ciegos sean dos pequeñas cámaras de fermentación, con una alta producción de ácidos grasos de cadena corta y vitaminas. Además, se describió la absorción de algunas hexosas y algunos aminoácidos en esta porción, provenientes de la fermentación microbiana.
| El ciego fue siempre el principal foco de estudios microbiológicos, ya que permite la proliferación de diversas cepas patogénicas, incluído el Clostridium perfringens. |
Los principales habitantes del ciego son las diferentes especies de Clostridios. Esta región es el único lugar de las aves donde es posible aprovechar parte de los carbohidratos estructurales de la dieta. Se ha demostrado que la capacidad de absorción de nutrientes en esta porción es significativa debido a que es posible relacionar la composición de la microbiota de los ciegos con el aprovechamiento de la energía de la dieta (Stanley et al., 2013).
Microhabitats
Las comunidades microbianas se establecen en diferentes microhabitats de una misma porción del tracto gastrointestinal. Este ecosistema puede ser dividido en dos componentes (Figura 5):
1) La comunidad dispersa en el lúmen
2) La comunidad adherida al mucus
La microbiota dispersa en el lúmen trae efectos importantes el el proceso de digestión del alimento. Las bacterias entran en contacto directo con las partículas del alimento, sus enzimas hidrolizan los compuestos, generando nutrientes para sí mismas y para el huésped; inclusive, varios de esos compuestos, influyen en la proliferación de las bacterias adheridas al mucus.
Figura 5. Microhabitats microbianos.
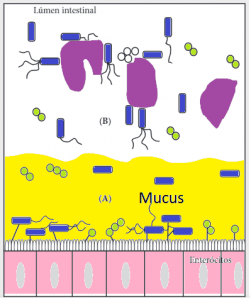

(A) Comunidad adherida al mucus, formado por bacterias ligadas al glucocálix o adheridas al mucus.
(B) Comunidad dispersa en el lúmen, formado por bacterias libres o adheridas a las partículas del alimento.
En este contexto, es posible entender la acción de varios probióticos, como Enterococcus y Bacillus que, por no poseer fimbrias, no se adhieren al epitelio, pero que hidrolizan compuestos que servirán de nutrientes para bacterias colonizadoras benéficas y también producen bacteriocinas que pueden evitar la proliferación de bacterias indeseables, como Salmonella y Clostridium.
La microbiota adherida al mucus es formada por bacterias extremamente adaptadas que se adhieren a la capa glicoproteica, una barrera física protectora natural del tracto. Esta adherencia específica facilita la colonización por comunidades deseables. El mucus está formado por una matriz proteica con una grand diversidad de oligosacáridos, que representan una fuente directa de carbohidratos y peptidos para las bacterias.
Esta simbiosis envuelve algunos mecanismos de señalización bacteria-huésped. Por ejemplo, los Lactobacilus secretan compuestos que, en las células epiteliales, estimulan la expresión de algunos genes responsables de la secreción de mucus. Por lo tanto, el huésped responde a la colonización bacteriana fortaleciendo sitios específicos de uniones y sustratos para su desarrollo, a cambio de una acción regulatoria que ejercen las bacterias contra los microorganismos patógenos. Es por eso, que en situaciones de desequilibrio de la flora normal, las cepas patógenas logran establecerse.
Es importante resaltar que la adherencia de bacterias comensales también causa una pequeña degradación celular de las vellosidades. Es por eso, que cuando las cepas benéficas están adheridas, esta pequeña degradación estimula la inmunidad local no específica, llevando a las aves a producir inmunoglobulinas que van a actuar en la mucosa intestinal. Esta acción ayudar a controlar la proliferación bacteriana excesiva de la flora normal y, consecuentemente, combatir las patógenas como Campylobacter♦
[/registrados]
Leer tambien Triptófano: impacto sobre la microbiota intestinal de los animales