



INTRODUCCIÓN
En la producción avícola, una estrategia ampliamente utilizada consiste en la administración de antibióticos en concentraciones subterapéuticas en la alimentación de las aves, conocidos como antibióticos promotores de crecimiento (APC) y se utilizan con el objetivo de estimular el crecimiento, mejorar la eficiencia de conversión de las aves y proteger la salud animal.
 |

La microbiota intestinal está involucrada en diversos procesos, como lo son la asimilación de los nutrientes de los alimentos, producción de vitaminas y aminoácidos esenciales y en la prevención de la colonización de microorganismos patógenos (Apajalahti, 2005).
La microbiota cecal en aves, además de estar involucrada en los procesos antes mencionados, participa en la fermentación de los productos de descomposición de polisacáridos no amiláceos (NSP), incluidos en la alimentación comercial de los pollos.
| La fermentación de los productos de descomposición de los NSP produce ácidos grasos volátiles de cadena corta (AGCC), que son absorbidos a través de la mucosa y catabolizados por el hospedador (McWhorter et al., 2009). Los AGCC, además de contribuir en la nutrición del pollo, ayudan a reducir el pH, lo que puede inhibir el crecimiento de patógenos (Apajalahti, 2005). |
Sin embargo, el uso de APC ha desarrollado preocupaciones sobre la aparición de cepas bacterianas resistentes a los antibióticos, que podrían comprometer la eficacia terapéutica de estos al ser empleados en medicina veterinaria y humana (Maron et al., 2013).
 |

Por lo tanto, es importante comprender los mecanismos involucrados en la acción de los APC, para desarrollar alternativas que simulen o asemejen los mecanismos de acción de los mismos.

MATERIALES Y MÉTODOS
El presente trabajo se llevó a cabo en la Sección Avicultura, INTA – EEA Pergamino, Argentina, utilizando 420 pollos machos de la línea Cobb-500 de un día de vida. Las aves fueron distribuidas formando lotes homogéneos y se alojaron en corrales a piso (1 m x 1,5 m), con viruta de madera de reuso como cama, 21 aves/corral. El alimento y el agua fueron provistos ad libitum.
|
Se utilizó un diseño de bloques completos al azar totalizando 2 tratamientos con 10 repeticiones cada uno (210 aves por tratamiento). El corral fue considerado la unidad experimental. Se evaluaron dos tratamientos, uno sin APC (tratamiento Basal) y el otro con Virginiamicina (VG) a 20 g/t como APC. |
Los pollos se criaron hasta los 42 días de edad y se determinó semanalmente el peso y el consumo, y se calculó la conversión y relación peso/conversión.








RESULTADOS
Parámetros zootécnicos
Las aves del tratamiento con VG presentaron un mayor peso que aquellas pertenecientes al tratamiento Basal, con diferencias significativas hasta los 27 días de edad (p≤0,05, Tabla 1).
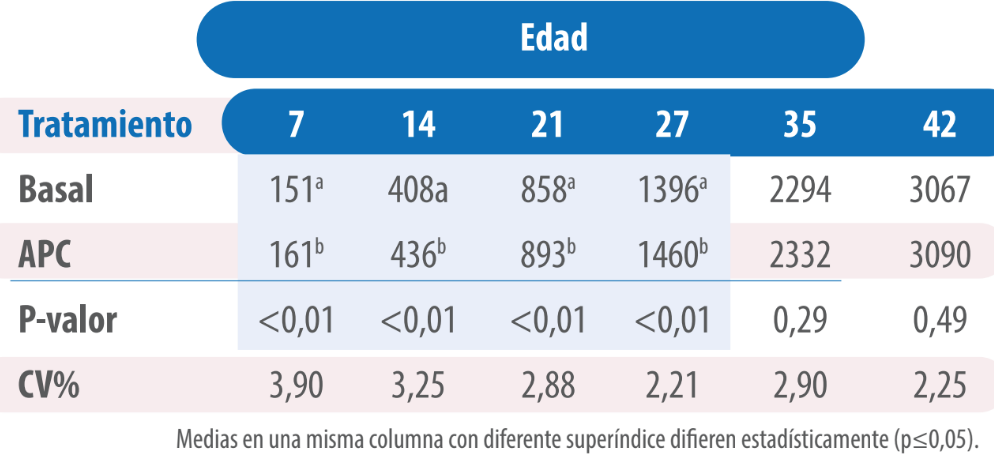

Tabla 1. Peso (g) de pollos alimentados con dietas sin y con virginiamicina
Asimismo, consumieron más alimento que aquellas pertenecientes al tratamiento Basal hasta los 27 días de edad, siendo esta diferencia estadísticamente significativa en las mediciones correspondientes a los días 7 y 14 días (p≤0,05, Tabla 2).
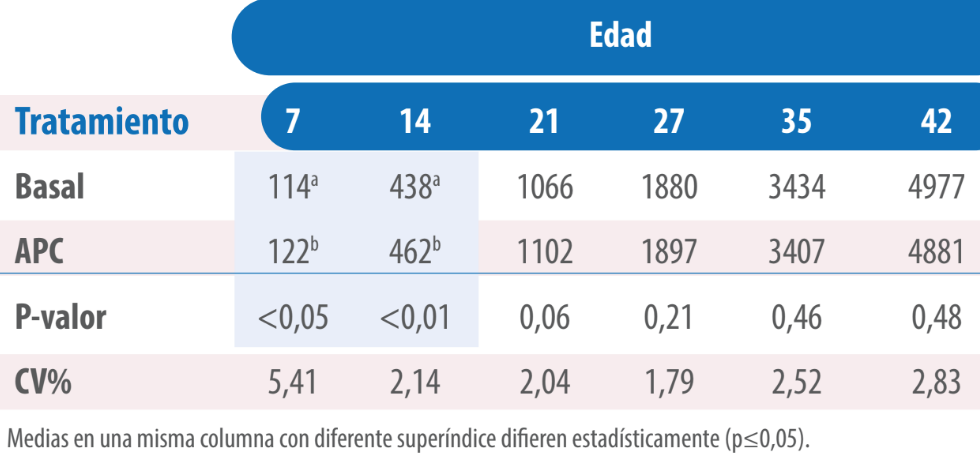

Tabla 2. Consumo (g) de pollos alimentados con dietas sin y con virginiamicina
Las aves del tratamiento con VG presentaron mejor conversión alimenticia que aquellas pertenecientes al tratamiento Basal, siendo esta diferencia significativa a los 21, 27, 35 y 42 días (p≤0,05, Tabla 3).
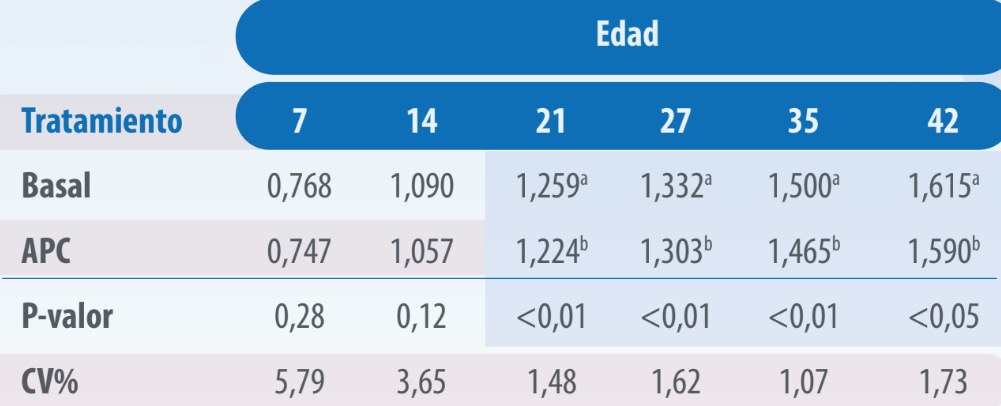

Tabla 3. Conversión de pollos alimentados con dietas sin y con virginiamicina
En cuanto a la relación peso/conversión, fue superior en las aves del tratamiento con VG que aquellas pertenecientes al tratamiento Basal, siendo esta diferencia significativa desde el inicio del ensayo hasta el día 35 (p≤0,05, Tabla 4).
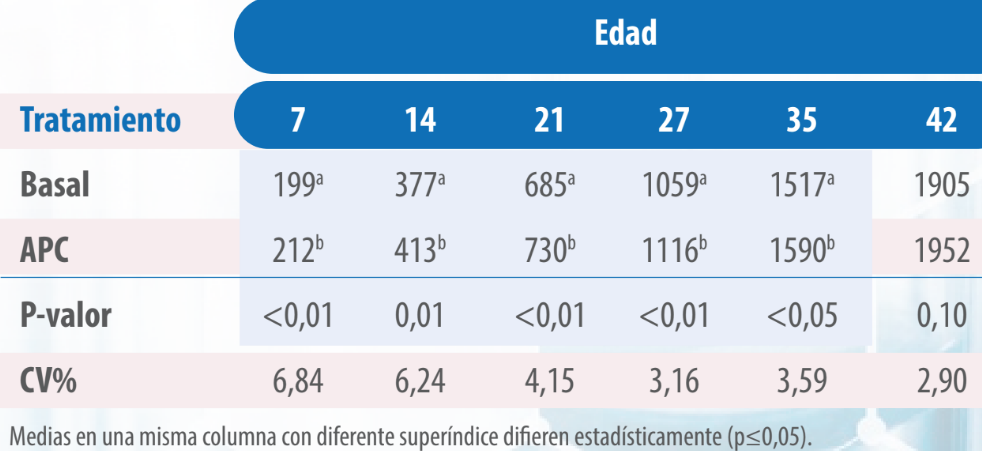

Tabla 4. Peso/conversión de pollos alimentados con dietas sin y con virginiamicina
En el análisis histomorfométrico del duodeno (Tabla 5) y del íleon (Tabla 6) se encontró que la relación vellosidad/cripta fue mayor en las aves tratadas con VG, siendo la diferencia significativa (p≤0,05) en las muestras tomadas a los 28 días (p≤0,05).
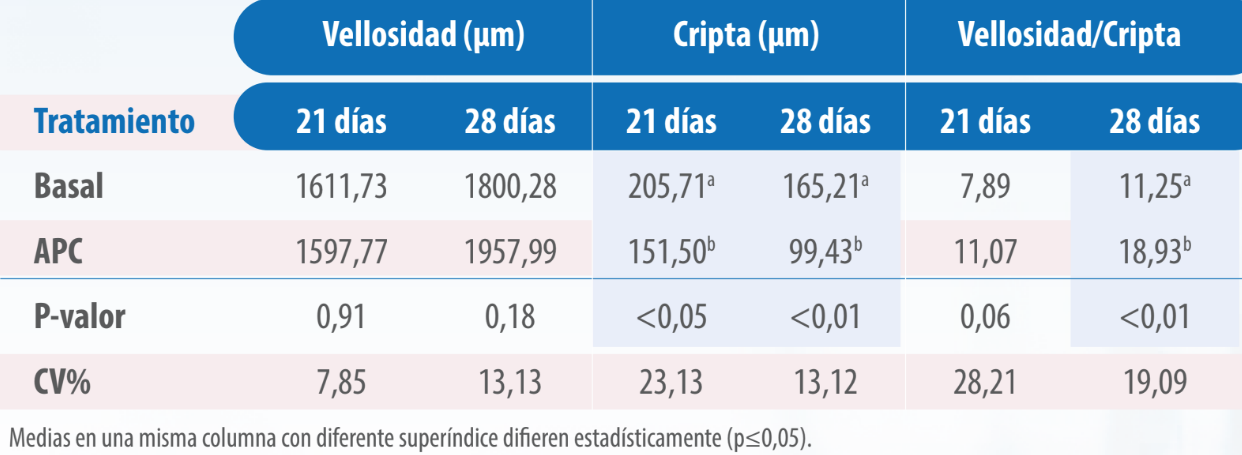

Tabla 5. Histomorfometría de duodeno de pollos alimentados con dietas sin y con virginiamicina
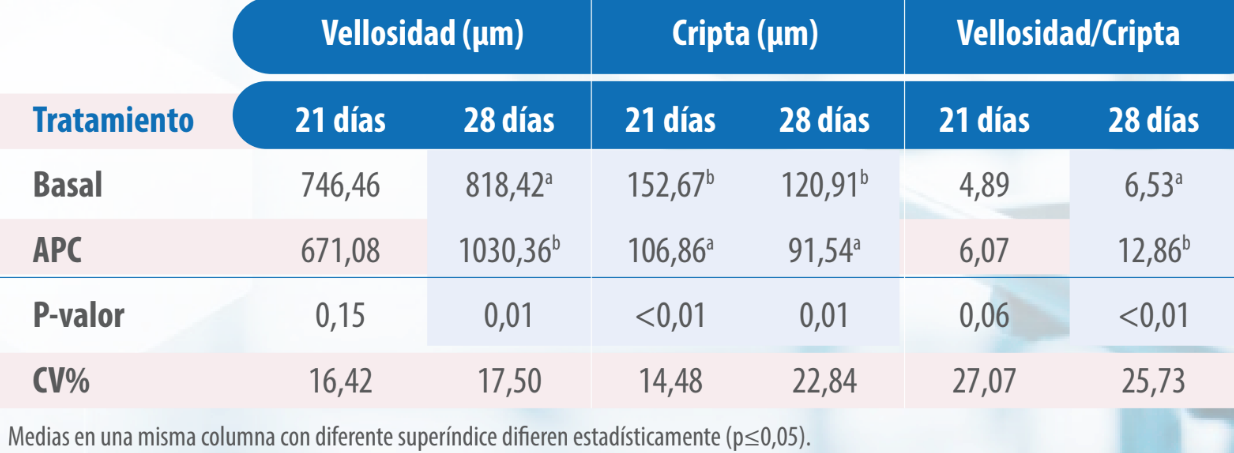

Tabla 6. Histomorfometría de íleon de pollos alimentados con dietas sin y con virginiamicina
Análisis de la composición de comunidades microbianas cecales


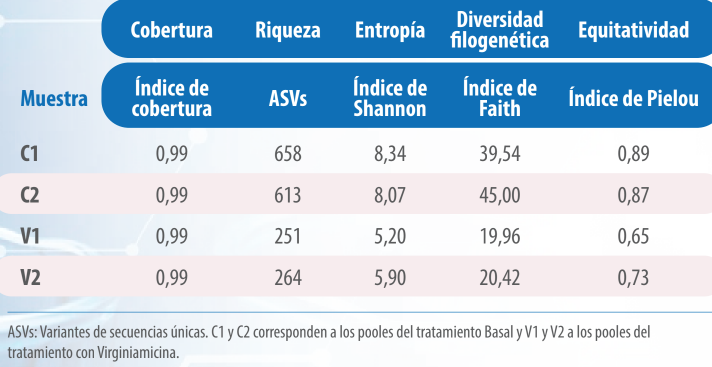

Tabla 7. Métricas de diversidad de pooles cecales de pollos de 21 días de vida.
Es importante señalar que, aunque se encontraron diferencias notables en la riqueza de especies en todas las métricas de diversidad analizadas, estas diferencias no alcanzaron significación estadística debido a la limitada cantidad de réplicas por tratamiento (p>0,05).
![]()



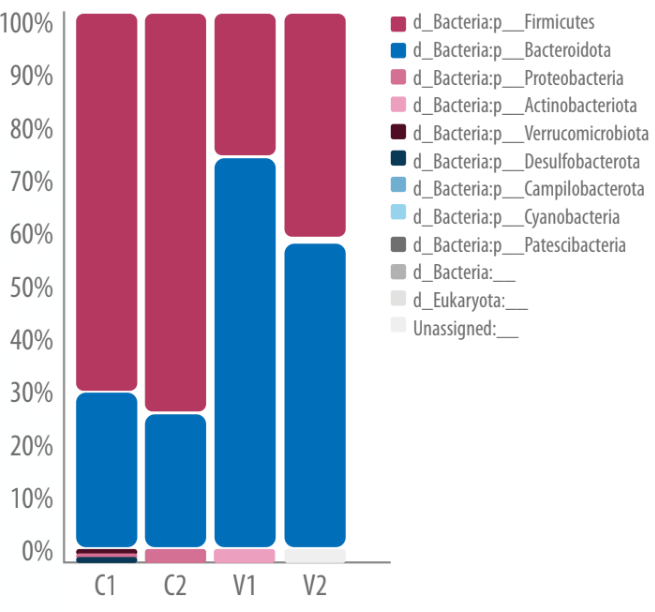

Figura 1. Perfiles de microbiota discriminados por muestra a nivel phylum.
C1 y C2 corresponden a los pooles del tratamiento Basal y V1 y V2 a los pooles del tratamiento con Virginiaminicina.
|
|

Correlación entre parámetros zootécnicos/histomorfométricos y microbiota
Los análisis de correlación de Spearman permitieron visualizar tendencias de asociación entre abundancias relativas de distintos taxones e indicadores relacionados con el rendimiento (Figura 2a) e histomorfometría (Figura 2b) de las aves.
Se encontraron 3 patrones bien diferenciados:
|
Bacteroides dorei y Alistipes, aumentan con VG y se asocian con una mejor productividad (mayor peso, menor consumo, menor conversión, mayor relación peso/conversión y mayor relación vellosidad/cripta); Eubacterium, que mostró un aumento con VG, pero no se identificó correlación con parámetros productivos; El resto de los taxones, disminuyen con VG y se asocian, en general, con menor eficiencia productiva (mayor consumo, menor peso, mayor conversión, menor peso/conversión, y menor relación vellosidad/cripta). |
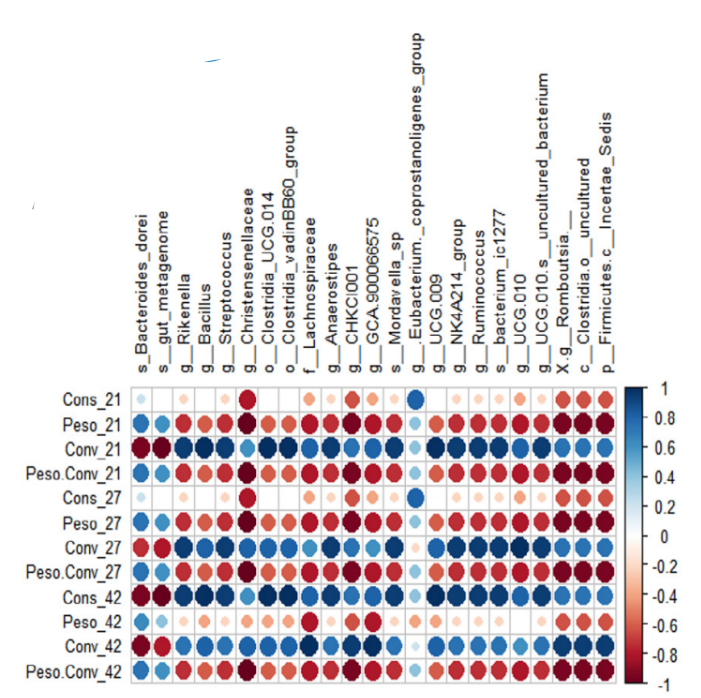

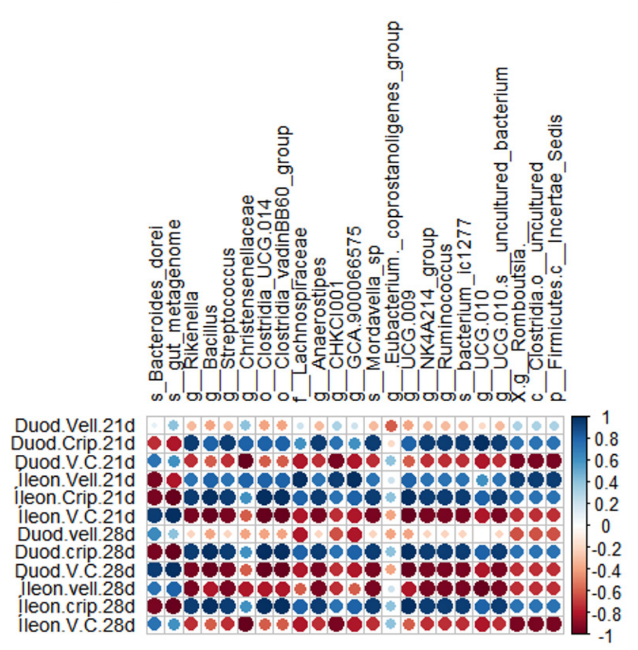

Figura 2. Gráficos de correlación de Spearman entre la abundancia relativa del microbioma intestinal y los indicadores relacionados con a) parámetros zootécnicos y b) parámetros histomorfométricos.
DISCUSIÓN
Ante las crecientes demandas de reducir o eliminar el uso de antimicrobianos en la cría de animales, resulta imperativo profundizar en la comprensión de los impactos generados por dichos agentes. De esta manera se podrán descubrir enfoques alternativos viables para salvaguardar la salud animal.
En el presente ensayo, la suplementación con VG produjo un mayor rendimiento de las aves en comparación con el tratamiento Basal, las cuales mostraron una mayor ganancia de peso en todas las edades, mayor eficiencia en la conversión y una mayor relación peso/conversión, lo que demuestra que la VG mejora la performance de las aves, coincidiendo estos resultados con los de otros autores (Ahmad et al., 2022; Engster et al.,2002).


Estos análisis brindan una explicación parcial de los motivos por los cuales las aves que reciben VG presentan un rendimiento superior en términos de eficiencia, por lo que se recurrió a la secuenciación de alto rendimiento de la región V3-V4 del gen ARNr 16S para analizar la composición bacteriana de la microbiota cecal.


Los índices de Shannon y de Pielou evidenciaron que la diversidad microbiana fue mayor y la distribución de las especies más equitativa en el tratamiento Basal y con mayor uniformidad en la composición microbiana de las muestras.
Estos resultados sugieren que el tratamiento con VG impacta en numerosas especies bacterianas y a su vez provoca un desbalance en la microbiota. Estas observaciones concuerdan con los hallazgos de Danzeisen et al. (2011).
![]()

En el presente estudio, el uso del APC tuvo un impacto significativo en la abundancia relativa de ciertos grupos taxonómicos en el ciego. Específicamente, se observó que 12 especies fueron completamente eliminadas debido a la presencia de VG, mientras que 9 taxones experimentaron una disminución notable en su abundancia.
Por otro lado, se registró un aumento significativo en la abundancia de 3 taxones en particular, entre los que se encontraron Alistipes, perteneciente a la familia Rikenellaceae; un género desconocido de la familia Eubacteriaceae y la especie Bacteroides dorei. A nivel phylum, se observó una disminución en la relación Firmicutes/Bacteroidetes en las aves tratadas con VG.
Según lo planteado por Dumonceaux et al. (2006), el aumento en la abundancia relativa de ciertas especies puede explicarse por una disminución inducida por VG en especies competidoras, lo cual, crea un espacio disponible para el crecimiento bacteriano.
![]()

![]()
| Esto sugiere que la abundancia relativa de estos taxones es mayor en los animales de mayor eficiencia productiva; entendiendo que dichos grupos bacterianos son relevantes para la productividad, y abren la posibilidad de hipotetizar e investigar acerca de su rol en la salud intestinal, la producción de AGCC y su interacción con otros miembros de la microbiota intestinal de las aves (Tabla 8). |
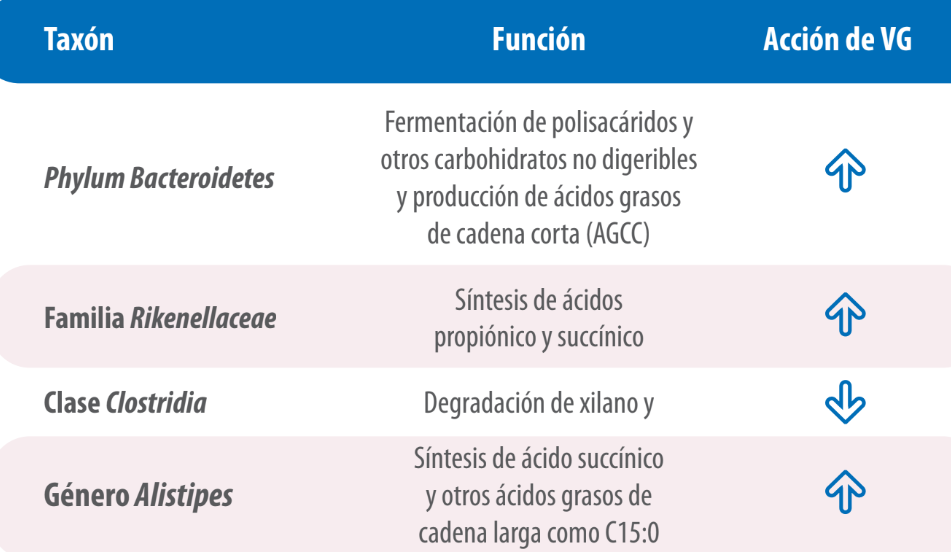

Tabla 8. Resumen de principales taxones, su función y la acción de virginiamicina sobre ellos
CONCLUSIÓN
|
|


BIBLIOGRAFÍA
Ahmad, M., Rasheed, M. A., Sattar, A., Abbas, G., & Basharat, A. (2022). Impact of antibiotic growth promoters (AGPs) in poultry production and alternative strategies. 1st International Conference on Sustainable Ecological Agriculture (1st ICSEA). Marzo 8-10. Konya, Turquía.
Apajalahti, J. (2005). Comparative Gut Microflora, Metabolic Challenges, and Potential Opportunities. Journal of Applied Poultry Research, 14(2), 444-453.
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., Al-Ghalith, G. A., Alexander, H., Alm, E. J., Arumugam, M., Asnicar, F., Bai, Y., Bisanz, J. E., Bittinger, K., Brejnrod, A., Brislawn, C. J., Brown, C. T., Callahan, B. J., Caraballo-Rodríguez, A. M., Chase, J., … Caporaso, J. G. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nature Biotechnology, 37(8), 852-857.
Chen, Y., Wang, J., Yu, L., Xu, T., & Zhu, N. (2020). Microbiota and metabolome responses in the cecum and serum of broiler chickens fed with plant essential oils or virginiamycin. Scientific Reports, 10(1), 5382.
Danzeisen, J. L., Kim, H. B., Isaacson, R. E., Tu, Z. J., & Johnson, T. J. (2011). Modulations of the chicken cecal microbiome and metagenome in response to anticoccidial and growth promoter treatment. PLoS ONE, 6(11), e27949.
Dibner, J. J., & Richards, J. D. (2005). Antibiotic growth promoters in agriculture: History and mode of action. Poultry Science, 84(4), 634-643.
Di Rienzo, J. A., Casanoves, F., Balzarini, M. G., González, L., Tablada, M., & Robledo, C. W. (2020). InfoStat v2020 [Software estadístico]. Córdoba, Argentina: FCA, Universidad Nacional de Córdoba.
Dumonceaux, T. J., Hill, J. E., Hemmingsen, S. M., & Van Kessel, A. G. (2006). Characterization of intestinal microbiota and response to dietary virginiamycin supplementation in the broiler chicken. Applied and Environmental Microbiology, 72(4), 2815-2823.
Engster, H. M., Marvil, D., & Stewart-Brown, B. (2002). The effect of withdrawing growth promoting antibiotics from broiler chickens: a long-term commercial industry study. Journal of Applied Poultry Research, 11(4), 431-436.
Klindworth, A., Pruesse, E., Schweer, T., Peplies, J., Quast, C., Horn, M., & Glöckner, F. O. (2013). Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Research, 41(1), e1-e1.
Maron, D. F., Smith, T. J., & Nachman, K. E. (2013). Restrictions on antimicrobial use in food animal production: An international regulatory and economic survey. Globalization and Health, 9(1), 48.
McWhorter, T. J., Caviedes-Vidal, E., & Karasov, W. H. (2009). The integration of digestion and osmoregulation in the avian gut. Biological Reviews, 84(4), 533-565.
Niewold, T. A. (2007). The nonantibiotic anti-inflammatory effect of antimicrobial growth promoters, the real mode of action? A hypothesis. Poultry Science, 86(4), 605-609.
Pourabedin, M., Xu, Z., Baurhoo, B., Chevaux, E., & Zhao, X. (2014). Effects of mannan oligosaccharide and virginiamycin on the cecal microbial community and intestinal morphology of chickens raised under suboptimal conditions. Canadian Journal of Microbiology, 60(5), 255-266.
Qureshi, S., Banday, M. T., Shakeel, I., Adil, S., Mir, M. S., Beigh, Y. A., & Amin, U. (2016). Histomorphological studies of broiler chicken fed diets supplemented with either raw or enzyme treated dandelion leaves and fenugreek seeds. Veterinary World, 9(3), 269-275.
Rahimi, S., Kathariou, S., Fletcher, O., & Grimes, J. L. (2019). Effect of a direct-fed microbial and prebiotic on performance and intestinal histomorophology of turkey poults challenged with Salmonella and Campylobacter. Poultry Science, 98(12), 6572-6578.
SENASA. (2015). Resolución 594 de 2015: Norma Técnica de Alimentos para Animales de la República Argentina. Recuperado de https://www.argentina. gob.ar/normativa/nacional/resoluci%C3%B3n-594-2015-256380/texto.
SENASA. (2018). Resolución 1119 de 2018: Modificación de la Norma Técnica de Alimentos para Animales de la República Argentina. Recuperado de https://www.boletinoficial.gob.ar/detalleAviso/primera/199463/20190103.