



Metabolismo de glucosa en Rumiantes
El metabolismo de glucosa en cualquier ser vivo es uno de los mecanismos más regulados del organismo.
El rumiante a diferencia de los monogástricos está preparado para sintetizar la mayor parte de la glucosa que necesita a partir del ácido propiónico (subproducto de la fermentación).
Figura 1: Recorrido del alimento en el sistema digestivo de un rumiante


Aunque el intestino delgado de los rumiantes está preparado para absorber glucosa, generalmente este carbohidrato no llega al intestino debido a que se fermenta rápidamente en el rumen (NRC, 2001).
En este proceso de fermentación ruminal, la glucosa se transforma en el ácido graso volátil de 3 carbonos: propiónico, el cual es absorbido por las paredes del rumen y transformado en glucosa en el hígado (Mc Donalds, 1995).
Figura 2: Proceso de la digestión de los carbohidratos en el sistema digestivo del rumiante
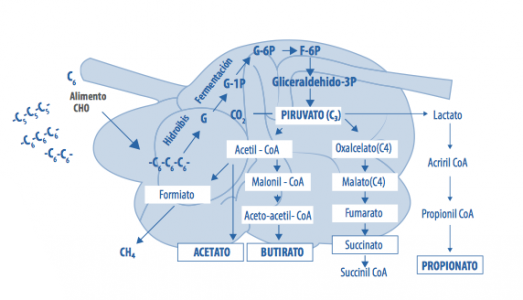

Metabolismo de almidón en rumiantes
El caso del almidón es distinto. Su metabolismo depende de:
- la dieta
- el tipo de animal que lo consume
- el tipo y método de procesamiento del grano que lo contenga
El almidón se degrada una parte en el rumen produciendo propiónico y otra parte termina siendo absorbido como glucosa en el intestino delgado o fermentado en el intestino grueso.
Owens y Zinn calculan que su digestibilidad total en el tracto digestivo en dietas de vacas de leche y de feedlot es de entre el 92 y el 98% (Owens y Zinn, 2005).
| Para poder calcular la energía aportada por el almidón como energía neta, es necesario conocer cuál es el sitio de digestión del mismo, donde es degradado y absorbido por el animal, en el rumen o en el intestino delgado (Harmon y McLeod 2001). |
El aporte de la fermentación del almidón por el intestino grueso es menor ya que el almidón que ha pasado por todo el animal hasta llegar al colon generalmente es menos degradable.
Digestión y absorción de la glucosa
Las relaciones estequiométricas teóricas y los experimentos de infusión controlada, claramente demuestran que la absorción de glucosa proveniente de almidón es más eficientemente convertida en tejidos cuando es absorbida a través del intestino delgado y no por la fermentación de la misma en el rumen.
Sin embargo, las limitaciones en la digestión del almidón y la absorción de glucosa por el intestino delgado muchas veces desperdician esta mayor eficiencia (Mac Leod, 2001).
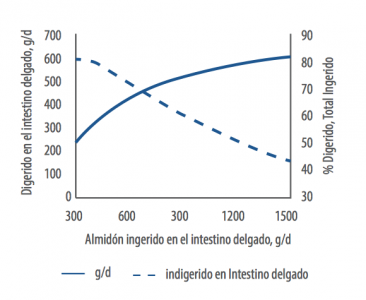
Huntington, en un intento de predecir la digestibilidad de almidón que entra en el intestino delgado, trazó una curva con rangos del 88% al 44% de digestibilidad cuando la cantidad de almidón que entra al intestino delgado fluctúa entre 300 y 1500gr (Huntington, 2006).

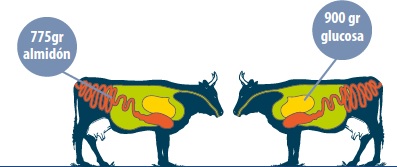
| Mc Leod en 2007, comparando el almidón con la glucosa como fuentes de energía, en uno de sus experimentos, infusionó 775 gr/día almidón hidrolizado y 900 gr/día glucosa, tanto en el intestino como en el rumen, a animales de 280 kg de peso vivo manteniendo una dieta basal.
↳ La infusión de glucosa provocó mayor tejido adiposo en los animales que la infusión de almidón. 1. El exceso de glucosa actúa como activador de lipogénesis en las células.
|

La glucosa necesita energía para ser absorbida ⇢ A diferencia de la glucosa la absorción de los ácidos grasos volátiles se da a través de las paredes del rumen sin un gasto de energía para el animal, la absorción de glucosa en cambio exige un gasto de energía para activar el sistema de transporte (GLUT 4) a través de la pared intestinal.
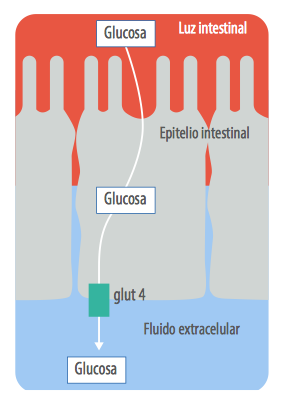

También conlleva un gasto de enzimas como la maltasa para degradar la maltosa en glucosa y luego absorberla (Baldwin, 1968 y 1995).
Por más que se gaste energía en la absorción de glucosa a través del intestino delgado no llega a compensar los 4 ATP de diferencia que exige convertir propionato a glucosa en el hígado con respecto a absorber glucosa directamente desde el intestino delgado (Mc Donalds 1995).
Cuando la glucosa es absorbida, los tejidos esplénicos y sobre todo la víscera portal tienen un doble efecto, por un lado no solo absorben sino que además consumen gran parte de lo absorbido, y por otro secretan hormonas como insulina o glucagón. A modo de ejemplo la víscera portal es una gran consumidora de glucosa, ácidos grasos volátiles y aminoácidos (Blouin, 2002).
Leer también “¿Qué carbohidratos encontramos en las células vegetales?”
Relación Energía con la retención de Nitrógeno
Glucosa en Carne y en Leche

[/registrados] La cantidad de glucosa absorbida en el intestino que no es consumida por los tejidos esplénicos y pasa al torrente sanguíneo para ser utilizados por músculo o glándula mamaria dependerá del tipo de animal analizado, del estado fisiológico del mismo y las condiciones ambientales a las cuales esté sometido.
En vacas de leche
El periodo de transición en la vaca de leche ha sido foco de muchos estudios estos últimos años, este periodo que aproximadamente involucra 21 días antes del parto y 28 días después del parto es objeto de cambios muy bruscos metabólicos y físicos para la vaca lechera de alto mérito genético Bell (1995) y Overton (2004).
Los estudios sugieren que la vaca experimenta una fuerte deficiencia de glucosa justo después del parto, esto sumado a el menor consumo de materia seca y a la gran demanda de nutrientes que exige la lactación gatilla distintos mecanismos metabólicos desarrollados por la vaca para sobrellevar esa falta de glucosa.
La activación de todos los mecanismos metabólicos para fabricar glucosa de distintos precursores como aminoácidos, glicerol, lactato y propionato es uno de ellos.


La resistencia a insulina es otra pero esta resistencia es selectiva ya que el feto y la glándula mamaria no lo son haciendo estos últimos tejidos los receptores de la mayor parte de la glucosa que se produzca o entre al organismo (Reynolds, 2001).
Después del parto…
Está bien demostrado que elevar la concentración de glucosa en sangre después del parto es fundamental para poder obtener buena reproducción, en su trabajo (Garverick et al., 2013) obtuvo mejor índice de preñez al primer servicio con vacas que presentaban mayores concentraciones de glucosa en sangre.
Animales con estrés por calor…
Algo similar ocurre con la vaca que sufre de stress por calor, en su trabajo Rhodes (2009) y Wheelock (2010) claramente demuestran que no toda la pérdida de producción se puede explicar por la falta de consumo de materia seca cuando la vaca sufre de stress por calor, ellos sugiere que solo el 50% de la pérdida en producción por stress por calor se explicaría por disminución en el consumo y que el 50% restante se explicaría por alteraciones metabólicas en la vaca.
Aparentemente la vaca que sufre de stress por calor está impedida de desmovilizar ácidos grasos no esterificados desde los tejidos grasos, no pudiéndolos usar como fuente de energía para compensar la falta de consumo.
| La posible explicación de la falta de movilización de tejidos grasos serían los altos valores de insulina encontrados en sangre ya que insulina es una potente hormona anti impolítica (O’Brien, 2010).
Esto sumado a la insensibilización a la insulina de casi todos los tejidos salvo la glándula mamaria y el feto presentaría una oportunidad para que la glucosa que fabrique el animal o ingiera pueda ser utilizada directamente para producir leche y preñarse. |
Russi et al. (2011) claramente demostró en su ensayo con vacas en transición que elevar los niveles de glucosa en sangre produjo más lactosa en leche y más condición corporal para las vacas que consumían un suplemento de glucosa protegida de la degradación ruminal. Utilizando el mismo suplemento, Mcarthy et al. (2020) detectaron menor concentración en sangre de ácidos grasos no esterificados, beta hidroxibuirato y marcadores inflamatorios como LPB y haptoglobina.
En animales de carne
Varios experimentos en terneros de la Universidad de Kansas reportan aumento de eficiencia de retención de aminoácidos y mejor uso de nitrógeno con el aumento del aporte de la concentración energética aportada (Schroeder 2006a, Schroeder 2006b, Awawadeh 2006).

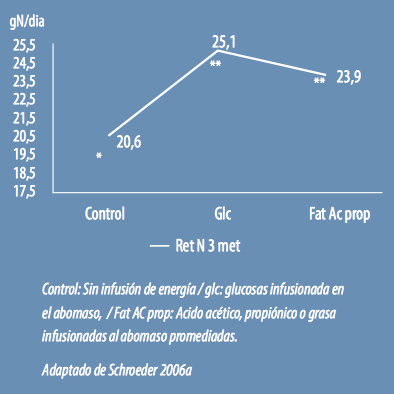
En uno de sus experimentos Schroeder (2006a) utilizo distintas fuentes de energía infusionadas o al abomaso o al rumen para evaluar la eficiencia de utilización de N y se infusionaron:

- en abomaso: glucosa a razón de 360 gr/día
- en rumen: acético, propiónico y butírico
Todos los excesos de energía respondieron haciendo más eficiente la utilización de nitrógeno sin diferencias significativas, pero numéricamente la glucosa fue la fuente de energía que produjo más eficiencia.
Eficiencia en el uso de nitrógeno con distintas fuentes de energía infusionadas en el abomaso

Russi et al. (2014) utilizó glucosa protegida en terneras de recría y observó que las que las terneras que consumían 180 gr de glucosa protegida durante todo el ciclo de engorde presentaban menos consumo y mejoraban la eficiencia de conversión medida como G:F.
No obstante el menor consumo por animal no se observaron diferencias en parámetros de concentración de glucosa en sangre o de insulina, ácidos grasos no esterificados (AGNES) o urea en plasma, sugiriendo que los animales que consumieron 180 gr de glucosa protegida tuvieron un metabolismo más eficiente.
Novillos en Terminación


En su experimento Mac Leod (2007) decidió infusionar almidón hidrolizado o glucosa tanto al intestino como al rumen, y una de las conclusiones de este ensayo fue que alimentar con glucosa infusionada en el intestino aumento la acreción de tejido adiposo en todo el animal pero particularmente en el tejido omental.
También se pueden encontrar ventajas en alimentar con glucosa protegida animales en terminación durante el verano, ya que aparentemente el mismo mecanismo de menor re movilización de tejido adiposo que actúa sobre la vaca estaría actuando en el animal de carne (O’Brien, 2010).
En su experimento Russi (2019) utilizó glucosa protegida como suplemento para alimentar novillos en terminación durante el verano donde el ITH fue de 72±4,9, la respuesta al suplemento se hizo más evidente en los picos de estrés por calor donde los animales alimentados con la glucosa consumieron menos pero engordaron lo mismo mostrando una eficiencia de conversión mejor.
En vacas de cría
En vacas de cría, Petersen et al. han encontrado que alimentar precursores gluconeogénicos ya sea como proteína pasante o como sales de propionato permite reducir los días al primer estro y % de preñez cambio en la condición corporal y peso del ternero al nacimiento en vacas de 2 o 3 años
Conclusión
Creemos que hay suficiente evidencia en la bibliografía para dejar de tratar a la energía como una caja negra y poder caracterizar las distintas fuentes con su particular eficiencia y modo de uso de acuerdo a los requerimientos del animal♦


[/registrados]