




Desempenho e nutrição de suínos em condição de desafios ambientais: estresse por calor e desafio sanitário

Em regiões de clima tropical e subtropical, os fatores climáticos merecem destaque uma vez que, nestas regiões, ocorre associação entre elevadas temperaturas e umidade relativa do ar. Quando expostos a condições de alta temperatura ambiental, suínos mantêm a homeotermia por meio de ajustes comportamentais, fisiológicos e metabólicos para diminuir a produção e/ou aumentar a dissipação de calor.
 Campos et al. (2014) observaram que suínos em crescimento quando expostos a condições de alta temperatura (30°C) apresentaram menor consumo de ração (-500 g/dia) e ganho de peso (-230 g/dia) quando comparados com suínos em condições de termoneutralidade (24°C).
Campos et al. (2014) observaram que suínos em crescimento quando expostos a condições de alta temperatura (30°C) apresentaram menor consumo de ração (-500 g/dia) e ganho de peso (-230 g/dia) quando comparados com suínos em condições de termoneutralidade (24°C).
Adicionalmente, Saraiva et al. (2011) relataram que animais em crescimento e terminação criados durante o verão na região sudeste do Brasil tiveram menor taxa de crescimento quando comparados com aqueles criados durante o inverno (910 vs. 1010 g/dia). No que diz respeito ao desempenho reprodutivo, o estresse por calor tem sido associado ao:
![]() Atraso da idade à puberdade
Atraso da idade à puberdade
![]() Maior intervalo desmame-estro
Maior intervalo desmame-estro
![]() Menor taxa de fecundidade e de concepção
Menor taxa de fecundidade e de concepção
![]() Maior incidência de leitões de baixo peso (Renaudeau et al, 2012; Campos et al., 2012)
Maior incidência de leitões de baixo peso (Renaudeau et al, 2012; Campos et al., 2012)
O genótipo de suínos também é um fator de grande importância uma vez que exerce influência sobre a capacidade de adaptação dos animais ao ambiente térmico.

![]() Em estudo com suínos submetidos a condição de estresse cíclico por calor, Fraga et al. (2019) observaram que animais com maior potencial para deposição de tecido magro (maior porcentagem de genes Pietrain) apresentam maior susceptibilidade a alta temperatura ambiental e, consequentemente, pior desempenho.
Em estudo com suínos submetidos a condição de estresse cíclico por calor, Fraga et al. (2019) observaram que animais com maior potencial para deposição de tecido magro (maior porcentagem de genes Pietrain) apresentam maior susceptibilidade a alta temperatura ambiental e, consequentemente, pior desempenho.
De fato, genótipos modernos tendem a ser mais susceptíveis ao estresse por calor devido à maior termogênese (Gaughan et al., 2009) e menor adaptabilidade a desafios ambientais.
Isso ocorre, devido a menor capacidade de mobilizar recursos fisiológicos e nutricionais para funções outras àquelas associadas ao crescimento e reprodução, tais como termorregulação e resposta imune (Rauw, 2012; Dumont et al., 2014).
![]() Por outro lado, a associação entre elevadas temperaturas ambientais e umidade relativa do ar favorece a proliferação e disseminação de vetores e/ou patógenos, resultando em maior pressão patogênica no ambiente (Campos et al., 2017) e com impacto significativo na produção animal uma vez que compromete o bem-estar, saúde e desempenho dos animais.
Por outro lado, a associação entre elevadas temperaturas ambientais e umidade relativa do ar favorece a proliferação e disseminação de vetores e/ou patógenos, resultando em maior pressão patogênica no ambiente (Campos et al., 2017) e com impacto significativo na produção animal uma vez que compromete o bem-estar, saúde e desempenho dos animais.
![]()
![]() Além disso, existe uma crescente demanda para reduzir a utilização de medicamentos de forma a permitir o desenvolvimento de sistemas sustentáveis de produção animal.
Além disso, existe uma crescente demanda para reduzir a utilização de medicamentos de forma a permitir o desenvolvimento de sistemas sustentáveis de produção animal.
![]() O uso de antimicrobianos promotores de crescimento na produção animal, por exemplo, foi associado a uma maior pressão de seleção de bactérias resistentes com sérios riscos para a saúde humana e animal (Pakpour et al., 2012).
O uso de antimicrobianos promotores de crescimento na produção animal, por exemplo, foi associado a uma maior pressão de seleção de bactérias resistentes com sérios riscos para a saúde humana e animal (Pakpour et al., 2012).
 Quando expostos a desafios sanitários (bactérias, vírus, parasitas, micotoxinas, más condições de higiene), mecanismos de defesa para manutenção da homeostase e integridade do organismo são ativados com consequências negativas para o desempenho produtivo e reprodutivo dos animais (Pastorelli et al., 2012).
Quando expostos a desafios sanitários (bactérias, vírus, parasitas, micotoxinas, más condições de higiene), mecanismos de defesa para manutenção da homeostase e integridade do organismo são ativados com consequências negativas para o desempenho produtivo e reprodutivo dos animais (Pastorelli et al., 2012).
Suínos respondem aos desafios sanitários por meio de ajustes neuroendócrinos e metabólicos na tentativa de restaurar a homeostase e integridade do organismo. Tais ajustes incluem:
![]() Febre
Febre
![]() Aumento da taxa metabólica para ativação e manutenção das respostas imunes e não imunes
Aumento da taxa metabólica para ativação e manutenção das respostas imunes e não imunes
![]() Redistribuição de nutrientes dos processos de crescimento e produção para a resposta imune
Redistribuição de nutrientes dos processos de crescimento e produção para a resposta imune
A febre é um dos sinais clínicos mais comuns em quadros de
infecção e inflamação, e o de maior custo energético.

Outras alterações metabólicas também são observadas em condição de desafio sanitário. Em estudo com suínos que receberam sucessivas aplicações de Escherichia coli (LPS), Escribano et al. (2014) observaram:
![]() Maior concentração salivar de haptoglobina
Maior concentração salivar de haptoglobina
![]() Proteína C reativa
Proteína C reativa
![]() Cortisol
Cortisol
![]() Imunoglobulina
Imunoglobulina
Quando comparado ao grupo controle (sem aplicação). O que indica ativação do sistema imunológico em condição de desafio sanitário.
![]() Similarmente, Fraga et al. (2021) observou aumento das concentrações plasmáticas de haptoglobina, neutrófilos e imunoglobulina G em suínos mantidos em más condições sanitárias de alojamento. Além dessas alterações metabólicas, a redução no consumo de alimentos (anorexia) também é uma resposta dos animais alojados em ambiente sanitariamente desafiador.
Similarmente, Fraga et al. (2021) observou aumento das concentrações plasmáticas de haptoglobina, neutrófilos e imunoglobulina G em suínos mantidos em más condições sanitárias de alojamento. Além dessas alterações metabólicas, a redução no consumo de alimentos (anorexia) também é uma resposta dos animais alojados em ambiente sanitariamente desafiador.
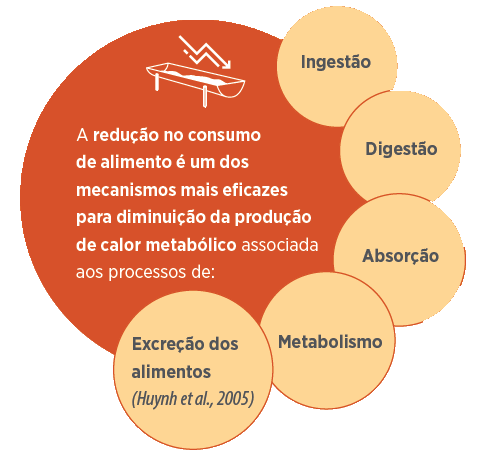
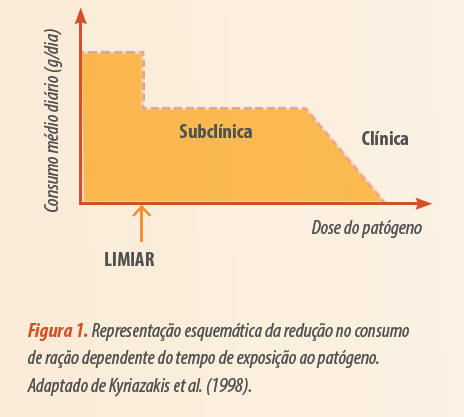
Mesmo a nível subclínico, cujas consequências não são visualmente observadas, Kyriazakis et al. (2008) observaram redução de 75-80% no consumo alimentar. Contudo, em uma condição de desafio mais intenso (tempo e/ou dose do patógeno), no qual sinais clínicos são observados, o consumo alimentar pode ser próximo de zero (Figura 1).
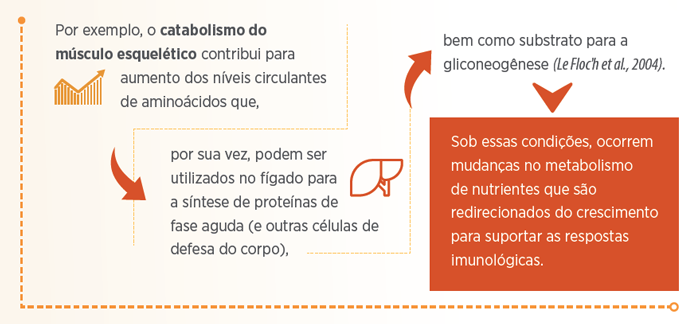
Uma vez que em condições de desafio sanitário os suínos podem se encontrar em estado anoréxico, são observadas alterações metabólicas (como lipólise do tecido adiposo e proteólise do músculo esquelético) para aumento da disponibilidade de nutrientes destinados a síntese de células imunes (Le Floc’h et al., 2004).
Sendo assim, Pastorelli et al. (2012) reportaram consistente diminuição no consumo alimentar e ganho de peso em suínos expostos e diferentes desafios sanitários. Por exemplo, foi relatada uma redução no consumo de ração de:
![]() 8% para infecções bacterianas digestivas
8% para infecções bacterianas digestivas
![]() 4% para condições precárias de higiene
4% para condições precárias de higiene
![]() 10% para inflamação induzida por LPS
10% para inflamação induzida por LPS
![]() 23% para micotoxicoses
23% para micotoxicoses
![]() 3% para infecções parasitárias
3% para infecções parasitárias
![]() 16% para doenças respiratórias.
16% para doenças respiratórias.
Dentre as estratégias para preservação da saúde e desempenho dos animais de produção submetidos a condições de desafio ambiental, crescente atenção tem sido voltada para a suplementação de aminoácidos.
Esses, são compostos orgânicos absorvidos e utilizados pelo organismo para síntese de proteínas, enzimas, hormônios, componentes estruturais, de transporte e do sistema imune. Aminoácidos atuam na:
![]() Manutenção da integridade e funcionalidade intestinal
Manutenção da integridade e funcionalidade intestinal
![]() Redução do estresse oxidativo
Redução do estresse oxidativo
![]() Processos de sinalização de resposta inflamatória e imune
Processos de sinalização de resposta inflamatória e imune
![]() Regulação das citocinas pró e anti-inflamatórias
Regulação das citocinas pró e anti-inflamatórias
(Liu et al., 2017; Le Floc’h et al., 2018).
| Potencial uso de estratégias nutricionais para suínos em diferentes condições de desafio |
 [registrados]
[registrados]
Em termos de nutrição animal, os efeitos do ambiente na fisiologia e metabolismo dos animais, e nas exigências nutricionais são raramente considerados. Majoritariamente, as dietas são formuladas em função da fase de crescimento dos animais (programa de alimentação por fases), independentemente das interações animal-ambiente.
![]() Para suínos em crescimento, por exemplo, uma única dieta é fornecida para animais entre 30 e 50 kg de peso vivo e outra para animais entre 50 e 70 kg de peso vivo (Rostagno et al., 2017). Além disso, nesses programas de alimentação, os nutrientes são geralmente fornecidos em excesso, a fim de suprir as exigências dos animais com maior demanda nutricional em um grupo de indivíduos.
Para suínos em crescimento, por exemplo, uma única dieta é fornecida para animais entre 30 e 50 kg de peso vivo e outra para animais entre 50 e 70 kg de peso vivo (Rostagno et al., 2017). Além disso, nesses programas de alimentação, os nutrientes são geralmente fornecidos em excesso, a fim de suprir as exigências dos animais com maior demanda nutricional em um grupo de indivíduos.
Essa prática, porém, conduz à formulação de dietas com alto valor termogênico e, portanto, inapropriadas para regiões de clima quente.
|
Dessa maneira, estratégias nutricionais podem ser utilizadas com o objetivo de melhor adequação das dietas frente às condições ambientais. Como forma de garantir o atendimento das exigências nutricionais em função da redução do consumo de alimento pelos animais, uma estratégia consiste no aumento dos níveis dos nutrientes (aumento da densidade nutricional) nas rações em condições de estresse por calor (Tavares, 2000). |
Essa estratégia pode ser realizada pela suplementação de energia por meio do uso de óleos e gorduras que, além de aumentar o valor energético da dieta, permite uma redução da produção de calor metabólico devido ao baixo incremento calórico desses ingredientes.
![]() Entretanto, é importante mencionar que existem limites para a adição de óleos e gorduras, uma vez que níveis acima do recomendado podem diminuir a digestibilidade dos nutrientes em virtude da menor ação das enzimas digestivas (Jansen et al., 2007) além de poder acarretar comprometimento na elaboração das rações devido a menor durabilidade dos pellets (Briggs et al., 1999).
Entretanto, é importante mencionar que existem limites para a adição de óleos e gorduras, uma vez que níveis acima do recomendado podem diminuir a digestibilidade dos nutrientes em virtude da menor ação das enzimas digestivas (Jansen et al., 2007) além de poder acarretar comprometimento na elaboração das rações devido a menor durabilidade dos pellets (Briggs et al., 1999).
 Outra estratégia consiste na formulação de dietas suplementadas com aminoácidos industriais com base no conceito de proteína ideal que, quando comparadas às dietas com base em proteína bruta, possuem baixo incremento calórico (Rodrigues et al., 2012; Fraga et al., 2019).
Outra estratégia consiste na formulação de dietas suplementadas com aminoácidos industriais com base no conceito de proteína ideal que, quando comparadas às dietas com base em proteína bruta, possuem baixo incremento calórico (Rodrigues et al., 2012; Fraga et al., 2019).
![]() Dietas formuladas com altos níveis de proteína bruta resultam em maiores taxas de transaminação, desaminação, síntese e excreção de ureia. Devido ao alto gasto energético dessas reações (ex: são necessários 4ATP para cada molécula de NH3 incorporada ao ciclo da uréia), o metabolismo proteico é considerado altamente calorigênico (van Milgen, 2001).
Dietas formuladas com altos níveis de proteína bruta resultam em maiores taxas de transaminação, desaminação, síntese e excreção de ureia. Devido ao alto gasto energético dessas reações (ex: são necessários 4ATP para cada molécula de NH3 incorporada ao ciclo da uréia), o metabolismo proteico é considerado altamente calorigênico (van Milgen, 2001).
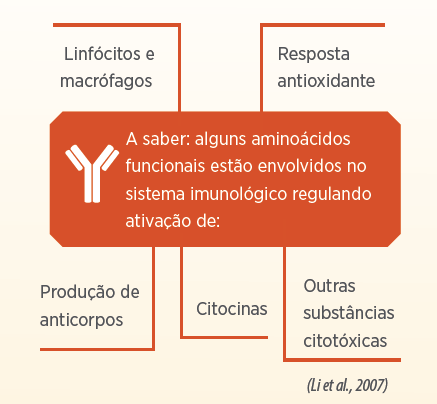
Especificamente, aminoácidos são nutrientes essenciais para manutenção do sistema de defesa (Le Floc’h et al., 2012). Aminoácidos funcionais são definidos como aqueles aminoácidos que regulam as principais vias metabólicas para melhoria da saúde, sobrevivência, crescimento, desenvolvimento, lactação e reprodução (Wu, 2010; Le Floc’h et al., 2018).
Em condições de desafio, parte dos nutrientes (aminoácidos) que seriam direcionados para o crescimento são redirecionados para a mecanismos de defesa e resposta imune.
 Estudos ressaltam a função imunometabólica da metionina (segundo aminoácido limitante para suínos), cujo metabolismo pode gerar antioxidantes intracelulares, como glutationa e taurina (Sun et al., 2016; Zhou et al., 2016). Adicionalmente, existem evidências que a estimulação do sistema imune afeta o metabolismo de triptofano.
Estudos ressaltam a função imunometabólica da metionina (segundo aminoácido limitante para suínos), cujo metabolismo pode gerar antioxidantes intracelulares, como glutationa e taurina (Sun et al., 2016; Zhou et al., 2016). Adicionalmente, existem evidências que a estimulação do sistema imune afeta o metabolismo de triptofano.
Em condição de sistema imune ativado, o interferon (IFNy) ativa a enzima indoleamina 2,3-dioxygenase (IDO) cuja função é o catabolismo do triptofano em quinurenina. Esta, por sua vez, está relacionada a funções essenciais de resposta imunológica (Le Floc’h et al., 2010; Le Floc’h et al., 2004).
Além disso, cerca de 10 a 13% das glicoproteínas presentes no muco é constituída pelo aminoácido treonina (Pluske et al., 2018). Percebe-se, portanto, que o status nutricional e o sistema imunológico estão interligados.
 Análise das respostas plasmáticas dos aminoácidos a diferentes desafios fornece algumas indicações sobre as mudanças no metabolismo dos aminoácidos quando a saúde está comprometida (Le Floc’h et al., 2018).
Análise das respostas plasmáticas dos aminoácidos a diferentes desafios fornece algumas indicações sobre as mudanças no metabolismo dos aminoácidos quando a saúde está comprometida (Le Floc’h et al., 2018).
![]() Por exemplo, menores concentrações plasmáticas de arginina foram observadas em suínos coinoculados com Mycoplasma hyopneumoniae e H1N1 vírus (Le Floc’h et al., 2014) para suportar a maior demanda para síntese de componentes imunológicos.
Por exemplo, menores concentrações plasmáticas de arginina foram observadas em suínos coinoculados com Mycoplasma hyopneumoniae e H1N1 vírus (Le Floc’h et al., 2014) para suportar a maior demanda para síntese de componentes imunológicos.
De fato, arginina é um aminoácido funcional que serve como precursor para a síntese de poliaminas, óxido nítrico (um potente mediador da resposta imune) e participa da síntese de outros aminoácidos, como creatina e ornitina, que estão envolvidos em importantes funções imunológicas (Li et al., 2007).
Adicionalmente, o estresse oxidativo, um mecanismo associado à resposta inflamatória, reduziu as concentrações plasmáticas de triptofano em leitões desmamados (Lv et al., 2012). Tais mudanças no metabolismo do triptofano induzidas pela inflamação estão associadas ao papel funcional desse aminoácido durante o processo inflamatório (Le Floc’h et al., 2018).
Em condições de estresse por calor, além da diminuição no consumo de alimento, é observado comprometimento da integridade e funcionalidade intestinal (Pearce et al., 2013). Consequentemente, ocorre diminuição na
absorção e disponibilidade de aminoácidos para os processos de mantença e produção.
Por exemplo, Morales et al (2016) reportaram diminuição nas concentrações plasmáticas de:
![]() Arginina (-28%)
Arginina (-28%)
![]() Histidina (-12%)
Histidina (-12%)
![]() Leucina (-10%)
Leucina (-10%)
![]() Lisina (-20%)
Lisina (-20%)
![]() Fenilalanina (-15%)
Fenilalanina (-15%)
![]() Treonina (-27%)
Treonina (-27%)
![]() Triptofano (-25%)
Triptofano (-25%)
![]() Valina (-17%)
Valina (-17%)
Em suínos expostos a condições de estresse cíclico por calor (24 a 38°C) quando comparado aos animais mantidos em condição de termoneutralidade (24°C).
Alterações como essas podem estar associadas ao efeito negativo do estresse por calor na expressão de transportadores intestinais de aminoácidos (Morales et al., 2014) e/ou maior demanda metabólica desses nutrientes para outras funções como proliferação celular, substrato energético, dentre outros (Morales et al., 2016).
| Considerações finais |
Estudos que consideram as interações entre saúde e nutrição de suínos em um contexto prático ainda é um potencial a ser explorado. A formulação de dietas que visam atender as necessidades nutricionais específicas conforme o estado fisiológico e de saúde do animal pode atenuar os efeitos negativos dos desafios ambientais, além de ser uma potencial alternativa ao uso de antimicrobianos e medicamentos.
Portanto, a suplementação de aminoácidos específicos pode contribuir para preservação do desenvolvimento, crescimento e saúde dos animais submetidos a episódios de desafio sanitário e estresse por calor.
Desempenho e nutrição de suínos em condição de desafios ambientais: estresse por calor e desafio sanitário
Alícia Zem Fraga
Departamento de Zootecnia, Universidade Estadual Paulista “Julio de Mesquita Filho” (UNESP, Jaboticabal)
Prof. Paulo Henrique Reis Furtado Campos
Departamento de Zootecnia, Universidade Federal de Viçosa
Bernabucci, U., N. Lacetera, L. H. Baumgard, R. P. Rhoads, B. Ronchi, and A. Nardone. 2010. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal: J. Anim. Biosci. 4: 1167-1183.
Briggs, J. L., D. E. Maier, B. A. Watkins, and K. C. Behnke. 1999. Effect of ingredients and processing parameters on pellet quality. Poult Sci. 78: 1464-1471.
Campos, P.H., Silva, B.A., Donzele, J.L., Oliveira, R.F., Knol, E.F., 2012. Effects of sow nutrition during gestation on within-litter birth weight variation: a review. Animal 6, 797-806.
Campos, P.H.R.F., E. Labussière, J. Hernández-García, S. Dubois, D. Renaudeau, J. Noblet. 2014. Effects of ambient temperature on energy and nitrogen utilization in lipopolysaccharide-challenged growing pigs. J. Anim. Sci. 92: 4909-4920.
Campos, P. H. R. F., N. Le Floc’h, J. Noblet, and D. Renaudeau. 2017. Physiological responses of growing pigs to high ambient temperature and/or inflammatory challenges. R. Bras. Zootec. 46:537-544.
Castro, I., L. Quisenberry, R. M. Calvo, M. J. Obregon, and J. Lado- Abeal 2013. Septic shock non-thyroidal illness syndrome causes hypothyroidism and conditions for reduced sensitivity to thyroid hormone. J. Mol. Endocrinol. 50:255-266.
Collin, A., Vaz, M.J., Le Dividich, J., 2002. Effects of high temperature on body temperature and hormonal adjustments in piglets. Reproduction, Nutrition, Development 42, 45-53.
Collier, R.J., Limesand, S.W., Rhoads, M.L., Rhoads, R.P., Baumgard, L.H., 2008. Homeorhesis during heat stress. Cabi, Wallingford, UK.
Dumont, B., González-García, E., Thomas, M., Fortun-Lamothe, L., Ducrot, C., Dourmad, J.Y., Tichit, M., 2014. Forty research issues for the redesign of animal production systems in the 21st century. Animal, 1-12.
Escribano, D., P.H. Campos, A. M. Gutiérrez, N. Le Floc’h, J. J. Cerón, e E. Merlot. 2014. Effect of repeated administration of lipopolysaccharide on inflammatory and stress markers in saliva of growing pigs. Vet. J. 200:393-397.
Fraga AZ, Campos PHRF, da Silva WC, Caetano RP, Veira AM, dos Santos LS, Hauschild L. 2019. Sequential feeding with high-fat/low-crude protein diets for two lines of growing-finishing pigs under daily cyclic high ambient temperature conditions. J. Anim. Sc.2019; 97: 2493–2504.
Fraga A. Z., Louveau I, Campos PHRF, Hauschild L, Le Floc’h N (2021) Selection for feed efficiency elicits different postprandial plasma metabolite profiles in response to poor hygiene of housing conditions in growing pigs. PLoS ONE16(3): e0246216.
Gaughan, J,B., Lacetera, N., Valtorta, S.E., Khalifa, H.H., Hahn, L., Mader, T., 2009. Response of domestic animals to climate challenges. In: Biometeorology of adaptation to climate variability and change. Ed. Ebi, K.L., Burton, I. and McGregor G.R. Springer Science, Heidelberg, Germany ,131–170.
Huynh, T.T.T., Aarnink, A.J.A., Verstegen, M.W.A., Gerrits, W.J.J., Heetkamp, M.J.W., Kemp, B., Canh, T.T., 2005. Effects of increasing temperatures on physiological changes in pigs at different relative humidities. J. Anim. Sci. 83:1385-1396.
Jansen, W.L., Cone, J.W., Geelen, S.N.J., Sloet van Oldruitenborgh-Oosterbaan, M.M., Van Gelder, A.H., Oude Elferink, S.J.W.H., Beynen, A.C. 2007. High fat intake by ponies reduces both apparent digestibility of dietary cellulose and cellulose fermentation by faeces and isolated caecal and colonic contents. Anim. Feed Sci. Technol. 33: 298-308.
Kyriazakis I., F. B., Sandberg, and W. Brindle. 2008. The prediction of the consequences of pathogen challenges on the performance of growing pigs. Mathematical Modelling in Animal Nutrition, 398-418.
Kyriazakis I., B. J. Tolkamp, and M. R. Hutchings.1998. Towards a functional explanation for the occurrence of anorexia during parasitic infections. Anim. Behav. 56:265-274.
Le Floc’h, N., C. Knudsen, T. Gidenne, L. Montagne, E. Merlot, and O. Zemb. 2014. Impact of feed restriction on health, digestion and faecal microbiota of growing pigs housed in good or poor hygiene conditions. Animal. 8:1632–1642.
Le Floc’h N., F. Gondret, J. J. Matte, and H. Quesnel. 2012. Towards amino acid recommendations for specific physiological and patho-physiological states in pigs. Proceedings of the Nutrition Society. 71: 425–432.
Le Floc’h, N., J. J. Matte, D. Melchior, J. Van Milgen, and Sève, B. 2010. A moderate inflammation caused by the deterioration of housing conditions modifies Trp metabolism but not Trp requirement for growth of post-weaned piglets. Animal. 4:1891-1898.
Le Floc’h, N., D. Melchior, and C. Obled. 2004. Modifications of protein and amino acid metabolism during inflammation and immune system activation. Livest. Prod. Sci. 87:37-45.
Le Floc’h N, Wessels A, Corrent E, Wu G, Bosi P. The relevance of functional amino acids to support the health of growing pigs. Anim Feed Sci Technol. 2018; 245: 104–116.
Li, P., Yin, Y.-L., Li, D., Woo Kim, S., Wu, G., 2007. Amino acids and immune function. British Journal of Nutrition. 98:237-252.
Liu, Y., Wang, X., Hou, Y., Yin, Y., Qiu, Y., Wu, G., Hu C.A., 2017. Roles of amino acids in preventing and treating intestinal diseases: recent studies with pig models. Amino Acids 49,1277-1291.
Lu, X-J., Q. Chen, Y.-J. Rong, F. Chen, e J. Chen. 2017. CXCR3.1 and CXCR3.2 differentially contribute to macrophage polarization in teleost fish. Journal of Immunology. 198:4692-4706.
Morales, A., F. H. Grageola, N. Garcia, B. Arce, B. Araiza, J. Yanez, and M. Cervantes. 2014. Performance, sérum amino acid concentrations and expression of selected genes in pair-fed growing pigs exposed to high ambiente temperatures. J. Anim. Physiol. Anim. Nutr. (Berl). 98:928–935.
Morales, A., S. E. M. Cota, N. O. Ibarra, N. Arce, J. K. Htoo, and M. Cervantes. 2016. Effect of heat stress on the serum concentrations of free amino acids and some of their metabolites in growing pigs. J. Anim. Sci. 94:2835-2842.
Netea, M.G., B. J. Kullberg, and J.W.M. Van der Meer. 2000. Circulating cytokines as mediators of fever. Arch. Clin. Infect. Dis. 31:S178-S184.
Nicholson, L.B. 2016. The immune system. Essays in Biochemistry. 60: 275–301.
Pastorelli, H., J. van Milgen, P. Lovatto, and L. Montagne. 2012. Meta-analysis of feed intake and growth responses of growing pigs after a sanitary challenge. Animal 6, 952-961.
Pearce, S. C., V. Mani, R. L. Boddicker, J. S. Johnson, T. E. Weber, J. W. Ross, R. P. Rhoads, L.H. Baumgard, and N. K. Gabler. 2013. Heat stress reduces intestinal barrier integrity and favors intestinal glucose transport in growing pigs. PLoS One 8:e70215.
Renaudeau, D., E. Huc, and J. Noblet. 2007. Acclimation to high ambient temperature in Large White and Caribbean Creole growing pigs. J. Anim. Sci. 85, 779-790.
Renaudeau D, Collin A, Yahav S, de Basilio V, Gourdine 489 JL, Collier RJ. Adaptation to hot climate and strategies to alleviate heat stress in livestock production. Animal 2012; 6: 707–728.
Rauw, W.M. 2012. Immune response from a resource allocation perspective. Frontiers in Genetics.
Riera Romo, M., D. Pérez-Martínez, e C. Castillo Ferrer. 2016. Innate immunity in vertebrates: an overview. Immunology. 148:125–139.
Rodrigues, N. E. B., Fialho, E. T., Zangeronimo, M. G., Cantarelli, V. S., Rodrigues, P. B., Filho, M. R., Filho, M. R., Gomides, E. M., Betarelli, R. P. 2012. Reduction in the protein level and addition of oil in diets for finishing pugs under different temperatures. Rev. Bras. de Zootec. 41: 1878-1883.
Rostagno, H.S. et al. 2011. Tabelas brasileiras para aves e suínos: composição de alimentos
e exigências nutricionais. 3.ed. Viçosa: UFV.
Sun, F., Y. Cao, C. Cai, S. Li, D. Yu, e Y. Yao. 2016. Regulation of nutritional metabolism in transition dairy cows: Energy homeostasis and health in response to post-ruminal choline and methionine. 11: e0160659.
Saraiva, A., Donzele, J.L., Oliveira, R.F., Abreu, M.L., Silva, F.C., Guimaraes, S.E., Kim, S.W., 2011. Phosphorus requirements for 60 to 100 kg pigs selected for high lean deposition under different thermal environments. J. Anim. Sci. 90 :1499-1505.
Pakpour S., S. Jabaji, e M. R. Chénier. 2012. Frequency of Antibiotic Resistance in a Swine Facility 2.5 Years After a Ban on Antibiotics. Microbial Ecology. 63:41-50.
Pluske, J., J. C. Kim, and J. L. Blank. 2018. Manipulating the immune system for pigs to optimise performance. Anim. Prod. Sci. 58: 666-680.
Spiers, D.E., 2012. Physiological basics of temperature regulation in domestic animals, In: Environmental Physiology of Livestock. Wiley-Blackwell Publications, Oxford, United Kingdom, pp. 17-33.
Tavares, S.L.S., Donzele, J.L., Donzele, R.F.M.O., Ferreira, A. S. 2000. Influência da temperatura ambiente sobre o desempenho e os parâmetros fisiológicos de suínos machos castrados dos 30 aos 60 kg. Rev. Bras. de Zootec. 29: 199-205.
van MILGEN, J. 2001. Nutritional in growing pigs: the animal, the diet and the environment. Factores que afectan la eficiéncia productiva y la calidad en el porcino. Institut de Recervca I Tecnologia Agroalimentàries.
Zhou, Z., M. Vailati-Riboni, E. Trevisi, J. K. Drackley, D. N. Luchini, e J. J. Loor. 2016. Better postpartal performance in dairy cows supplemented with rumen-protected methionine compared with choline during the peripartal period. J. Dairy Sci. 99: 8716–8732.
Wu, G., 2010. Functional amino acids in growth, reproduction and health. Adv. Nutr. 1, 31–37.
[/registrados]








