



En cualquier especie animal, la modulación del comportamiento ingestivo implica cambios en su patrón de consumo a corto plazo -distribución, número, tamaño y duración de las comidas a lo largo del día- partiendo de la base de que el consumo voluntario fluctúa influido por los mecanismos fisiológicos que lo regulan (Nelson, 1999; Howie et al., 2009).
Para regular el consumo voluntario el cerebro debe modular el apetito y dicha regulación ocurre a dos niveles: central y eje intestino-cerebro
Regulación del comportamiento ingestivo – Vías hedónica & homeostática
La principal regulación del comportamiento ingestivo se sitúa a nivel del sistema nervioso central, donde el cerebro recibe señales hormonales, nerviosas y metabólicas que regulan el estado energético del animal.
En respuesta a estas señales, se coordinan cambios en el consumo y liberación de energía (Cummings and Overduin, 2007).
Estos niveles de regulación se corresponden con la vía hedónica o de recompensa (Berridge, 2009) y la vía homeostática (Berthoud, 2011), respectivamente.
Ambas vías, hedónica y homeostática, actúan de forma interrelacionada por lo que conviene considerarlas en conjunto al momento de evaluar estrategias que tengan como objetivo principal modular el comportamiento ingestivo (Lutter y Nestler, 2009; Tulloch et al., 2014).


En este caso, la respuesta coincide con un aumento en la concentración sanguínea de hormonas causantes de saciedad, lo cual eventualmente resulta en una alteración del patrón de consumo del individuo y una disminución de la cantidad consumida
de dicho alimento (Stenblom et al., 2015).
» Vía hedónica
La vía hedónica se refiere a la activación (o inhibición) de los centros del placer, es decir, de los sistemas de recompensa situados en el cerebro cuya estimulación promueve el consumo independientemente del estado energético del animal.
Esta vía consta de dos componentes:
- Anticipación a la respuesta de placer
- Reacción placentera propiamente dicha que resulta de la experiencia sensorial ocurrida (Finlayson et al., 2007).
El gusto es la característica sensorial de los alimentos aparentemente más fácil de categorizar como hedónico o no hedónico


⇒ Gusto y olfato . Características sensoriales
Determinadas características sensoriales del alimento (ej., compuesto detectados por los sentidos del gusto y olfato) son capaces de estimular la vía hedónica al ser percibidos por el animal (Roura et al., 2008; Ginane et al., 2011; Clouard et al., 2012).
Además, en todas las especies, con mayor o menor habilidad, los individuos asocian dichas características con las consecuencias buenas o malas derivadas de su consumo, lo que se conoce como consecuencias post-ingestivas.
Más aún, las características sensoriales gustativas y olfativas de los alimentos intervienen en los procesos de aprendizaje asociativo y/o social, e incluso aquellos que operan antes del nacimiento facilitando el aprendizaje prenatal o “imprinting” (Smitzis et al., 2008; Oostindjer et al., 2010, 2011; Favreau et al., 2010; Figueroa, 2012; Clouard et al., 2014; Guzmán, 2014).
Además de estar estrechamente relacionado con el tipo de información que recibe el animal relativo al contenido nutritivo de la dieta, el gusto es la característica sensorial de los alimentos aparentemente más fácil de categorizar como hedónico o no hedónico en las distintas especies (Tabla 1 – página siguiente).
Eventualmente, esto dispara una cascada de eventos a nivel cerebral que culmina con la liberación en los centros de placer de neuropéptidos, tales como la dopamina, opioides, y canabinoides (Cota et al., 2006; Berridgeet al., 2009; Lutter y Nestler, 2009; Tedó, 2009; Favreau-Peigné et al., 2013).
Tipos de gustos básicos
Dentro de los gustos básicos reconocidos se encuentra el dulce, umami, amargo, salado y ácido, y recientemente se ha propuesto incorporar a dicha lista el gusto de ácidos grasos (da Silva et al., 2014; Liman et al., 2014).
El dulce y el umami, los cuales ayudan en la identificación de fuentes de azúcares y aminoácidos respectivamente, son considerados gustos hedónicos para la mayoría de los animales de granja (Roura y Tedó, 2009; Ginane et al., 2011; Roura et al., 2013).
Una excepción a esta observación pueden ser los pollos, los cuales no poseen receptor para detectar el gusto dulce y por tanto cabría esperar que no haya respuesta alguna a la estimulación dulce en esta especie (Roura et al., 2013).
Por otro lado, el gusto amargo es generalmente aversivo (Favreau et al., 2010), a pesar de que la respuesta depende también de la especie en consideración. Por ejemplo, en el caso de los rumiantes lo caprinos presentan una mayor aceptación o tolerancia a sustancias amargas que los bovinos (Ginane et al., 2011).
El mecanismo de acción de la vía hedónica involucra la interacción de los compuestos químicos del alimento -ej., azúcares, aminoácidos, minerales- con los receptores gustativos presentes en la cavidad oral


Tabla 1. Clasificación de la percepción sensorial gustativa como hedónica (aceptación – preferencia) o no hedónica (indiferencia – rechazo) en diferentes animales de abasto.
1 Fuentes: Roura y Tedó, 2009 (porcino), Ginane et al., 2011 (rumiantes), Roura et al.2013(aves) 2 En general, mayor aceptabilidad y preferencia en animales jóvenes o adultos en periodo de cría
3 Alta preferencia y aceptabilidad por el gusto umami en ovino independientemente de la edad 4 Mayor tolerancia a compuestos amargos en el ganado caprino 5 Efectos postingestivos
Comportamiento ingestivo1
De los tres, el sistema opioide parece ser el que juega un papel más relevante en la regulación del comportamiento ingestivo en respuesta a la percepción sensorial del alimento (Barbano y Cador, 2006; Montoro et al., 2012).
En términos de comportamiento ingestivo, la estimulación del sistema opioide resulta en una mayor aceptación de la dieta que contiene las características sensoriales que lo estimulan, una preferencia clara por la misma cuando se le da la opción al animal de escoger entre otras y un refuerzo de su consumo cuando va ligado a consecuencias postingestivas positivas para el individuo (Figura 1).
Sin embargo, en situaciones estresantes para el animal puede ocurrir que la respuesta a la estimulación hedónica sea menos intensa o más variable entre individuos. Este fenómeno se conoce como anhedonia inducida por estrés (Tanja y Epel, 2007; Torres y Nowson, 2007; Korte et al., 2009).
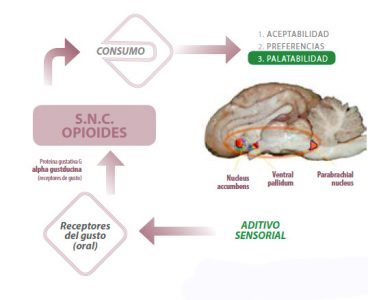

Figura 1 Esquema del mecanismo de acción de aditivos sensoriales con actividad en los centros hedónicos (opioides) a nivel del sistema nervioso central (S.N.C.) y su efecto sobre el comportamiento ingestivo
» Vía homeostática
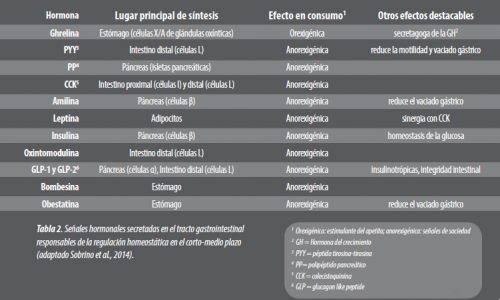
En la vía homeostática, se regula el consumo voluntario mediante las señales de saciedad y de apetito, cuya principal función consiste en optimizar la digestión y la absorción de nutrientes. Dicha modulación puede diferenciarse en una regulación de corto o largo plazo (Cummings y Overduin, 2007; Sobrino et al. 2014).
En estrategias de modulación de consumo en animales de producción gana especial interés la regulación en el corto plazo, la cual influye en la motilidad gastrointestinal y en la secreción de péptidos (hormonas) a lo largo del día, y por tanto, tiene un efecto directo sobre el patrón de consumo, productividad y bienestar animal a diario (Tabla 2).



Desde la perspectiva sensorial, se ha avanzado en el conocimiento de la regulación del consumo voluntario por la vía homeostática.
La estimulación de sensores localizados en el tracto gastrointestinal, los cuales son idénticos a los receptores gustativos que se localizan a nivel oral, poseen como función principal la detección de nutrientes disponibles (Geraedts, 2010; Liou, 2013; Steinert et al., 2013).
Estos sensores nutritivos son capaces de regular procesos fisiológicos y metabólicos, tales como el consumo voluntario, la motilidad intestinal, la digestión y la absorción de nutrientes, y por tanto, abren la posibilidad a potenciales aplicaciones sensoriales destinadas a mejorar aspectos productivos (Liu et al., 2013)
Estrategia para estimular la ingestión
Una estrategia interesante para estimular el comportamiento ingestivo a través de la vía homeostática en animales de abasto sería el aumentar la secreción de ghrelina endógena.
La ghrelina es una hormona de entre 26 y 28 aminoácidos, dependiendo de la especie, producida en distintas partes del tracto gastrointestinal, aunque mayormente se da su producción en el estómago en cerdos, abomaso en rumiantes y proventrículo en pollos y pavos.
En todas estas especies, se define como la hormona estimulante del apetito, por tanto orexigénica, e inductora de la secreción de la hormona del crecimiento, de gran relevancia, dado que influye en mejoras no tan solo asociadas al consumo voluntario (Salfen et al., 2004; Richards et al., 2006; ThidarMyint et al., 2006; Kaiya et al., 2007).
Para tener actividad biológica, la ghrelina precisa una modificación enzimática de acilación, lo cual ocurre en tan solo un 10-20% del total de ghrelina circulante. Por este motivo cuanto mayor sea la secreción de ghrelina total, cabría esperar una mayor cantidad de ghrelina activada (Kojima y Kangawa, 2005; Janssen et al., 2011).
En estudios con lechones y terneras, en los que se les ha inyectado ghrelina exógena de manera experimental, han sido demostrados los efectos positivos anteriormente descritos, como mejoras en el consumo y en el crecimiento de los animales tratados (Salfen et al., 2004; ThidarMyint et al., 2006).
Estrategia para aumentar la sensación de saciedad
Si por el contrario, el objetivo consistiera en aumentar la sensación de saciedad, podría utilizarse el uso de estrategias nutricionales, como:
- Inclusión de determinadas fuentes de fibra en la dieta para aprovechar sus efectos mecánicos de llenado y/o a aquellos derivados de su fermentación, como es la producción de ácidos grasos de cadena corta (Da Souza, 2013; Lindberg, 2014).
- Alternativamente, se podrían usar estrategias sensoriales específicas, distintas de los originadas por los productos de fermentación, que desencadenasen la secreción de señales de saciedad a nivel del intestino delgado, o incluso la modulación del tránsito gastrointestinal, como ocurre con el uso de sustancias amargas o pungentes (Reinbach, 2008; Janssen et al., 2011; Stenblom et al., 2015).


Figura 2. Esquema del mecanismo de acción de aditivos sensoriales con actividad homeostática (secreción de péptidos a lo largo del tracto gastrointestinal) y su efecto sobre el comportamiento ingestivo.
En el caso de la estimulación de la secreción de ghrelina o señales anorexigénicas, el mecanismo de acción que daría lugar a ello por medio de la percepción sensorial a nivel postoral involucraría la activación de sensores gustativos que se encuentran presentes en el tracto gastrointestinal, desde el estómago hasta el colon (Geraedts, 2010; Janssen et al., 2011; Liou, 2013; Steinert et al., 2013), los cuales utilizan los mismos mecanismos de transducción que en los receptores gustativos orales, y donde la proteína α- Gustducina, o α-Transducina, jugaría un papel relevante en la actividad de estos sensores, así como también en la modulación de la actividad de las hormonas secretadas (Figura 2).
Por tanto, la respuesta ingestiva a esperar con el uso de estrategias sensoriales que utilicen la vía de regulación homeostática, será una modulación del patrón de consumo diario que consistirá en un aumento del apetito (ghrelina) o en su inhibición (señales de saciedad), llegando al final a originar una respuesta de mayor o menor consumo en el animal.
Usos potenciales de la modulación del comportamiento ingestivo para mejorar la productividad y el bienestar animal
En este sentido, nuestro grupo ha trabajado ampliamente sobre el uso de estrategias sensoriales que pueden influir a la vez en la regulación de ambos mecanismos de modulación del comportamiento ingestivo -homeostático y hedónico- (Villalba et al., 2011; Montoro et al., 2012); y en cómo esta modulación es capaz de generar beneficios, tanto productivos como de bienestar animal, en distintas especies y en diferentes estadios productivos (Bargo et al., 2014; Mereu et al., 2013; Merrill, 2013; Montoro et al., 2011; Tedó et al., 2011, 2012).
Las aplicaciones prácticas donde tiene sentido pensar en la modulación del consumo voluntario como estrategia son diversas y específicas a la especie, donde deben combinarse ambos mecanismos de acción -homeostático y hedónico- para compensar las diferencias individuales y para obtener respuestas más uniformes.
Estas diferencias individuales suelen ser acentuadas por situaciones de estrés comúnmente presentes en nuestros sistemas productivos, tales como el destete precoz, el estrés térmico, los cambios de alimentación, la mezcla de individuos o el transporte.
Las situaciones de estrés pueden originar desde una respuesta de anhedonia (o pérdida de la capacidad de sentir placer) a una disminución del consumo voluntario derivado de la propia activación del eje hipotálamo- hipófisis –adrenal.
Las estrategias sensoriales basadas en conseguir una adaptación-familiarización previa, jugarán un papel de gran relevancia sobre el consumo; como también en otros aspectos, tales como productivos y de bienestar, incluyendo el garantizar una salud digestiva durante la vida productiva del animal (www.fawec.org).
Para más información, contactar con el equipo de ZOOTECNIA de Lucta – web LUCTA- División Zootecnia