


En los últimos números de nutriNews hemos dado un repaso sobre el metabolismo ruminal y sus procesos; desde los nutrientes de entrada con la dieta, pasando por sus reacciones en un medio de fermentación anaerobia, hasta los productos finales del metabolismo ruminal.
En este último artículo de la serie hablaremos sobre el metabolismo de los ácidos grasos volátiles (AGV) o ácidos grasos de cadena corta (AGCC).
DIGESTIÓN RUMINAL
En el ambiente o ecosistema ruminal, se genera energía y proteína.
Más del 60% de la energía que el animal utiliza para mantenimiento y producción (crecimiento, lactación, gestación, lana, etc.) procede de los ácidos grasos volátiles (AGV), y entre el 60 y el 80% de la proteína necesaria para el crecimiento y producción, proviene de la población microbiana del retículo-rumen.
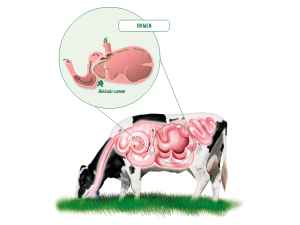

Siendo así, el rumen se presenta como un segmento del tracto digestivo clave para la supervivencia en condiciones extremas y para un óptimo rendimiento productivo.
De este modo , en la medida en que se mejoren los procesos de la digestión de los alimentos en el rumen, se mejorará sustancialmente la producción animal y con ella, la productividad del sistema ganadero.
CARBOHIDRATOS FIBROSOS
Todos los alimentos (forrajes o concentrados) están constituidos por distintas fracciones: carbohidratos, proteínas, lípidos, vitaminas, minerales y agua, de estos son los carbohidratos la fuente más importante de energía y los principales precursores de grasa y azúcar (lactosa) de la leche.
Los microorganismos del rumen permiten obtener energía de los carbohidratos fibrosos (celulosa y hemicelulosa):
la fibra en general, es voluminosa y queda retenida en el rumen en donde se fermenta lentamente y sirve de sostén para los microorganismos (MOO) ruminales.
Existen diferentes calidades de fibras en función del tipo y el lugar de crecimiento del vegetal y la edad de las plantas.
Al madurar la planta, el contenido de lignina de la fibra incrementa y el grado de fermentación de la celulosa y la hemicelulosa en el rumen se reduce.
El proceso de lignificación de las plantas no sólo se debe a la edad también depende del clima cuanto más calor más lignificación. La presencia de la fibra en las partículas largas es necesaria para estimular la rumia.
La rumia aumenta la separación y fermentación de la fibra, estimula las contracciones del rumen y aumenta el flujo de saliva hacia el rumen.
La saliva contiene bicarbonato de sodio y fosfatos que ayudan a mantener la acidez (pH) del contenido del rumen en un pH cercano a la neutralidad.
CARBOHIDRATOS NO-FIBROSOS
Los carbohidratos no-fibrosos (almidones y azúcares) fermentan en forma rápida y en gran porcentaje en el rumen.
El contenido de carbohidratos no-fibrosos incrementa la densidad de energía en la dieta, mejora el suministro de energía y determina la cantidad de proteína bacteriana producida en el rumen.
Sin embargo, los carbohidratos no-fibrosos no estimulan la rumia o la producción de saliva y cuando se encuentran en exceso pueden inhibir la fermentación de fibra.
![]()

FERMENTACIÓN RUMINAL
PROPIONATO
La digestión ruminal del almidón, genera una alta producción de AGV, destacándose el propionato (C3) cuya proporción molar aumenta con respecto a la fermentación ruminal del forraje fibroso, donde se genera una mayor proporción molar de acetato (C2).
El propionato presente en el rumen se absorbe por las paredes (más del 80 %) llegando al hígado, donde a través de un proceso metabólico (gluconeogénesis) se transforma en glucosa.
ACETATO Y BUTIRATO
Mientras que el acetato y butirato generan directamente energía en forma de ATP en rumen (Van Huotert, 1993).
En los animales de alta producción, el rumen, el hígado y la glándula mamaria son los principales órganos involucrados en el metabolismo de los carbohidratos.
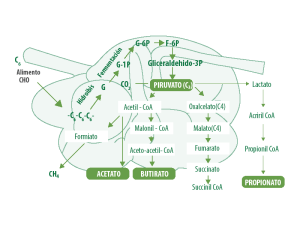

Durante la fermentación ruminal, la población de microorganismos, principalmente bacteriana, fermenta los carbohidratos para producir energía, gases (metano – CH₄ y bióxido de carbono – CO₂), calor y ácidos.
Las vías terminales que intervienen en la formación de los productos finales de la fermentación de los carbohidratos son muy complejas, son distintos mecanismos de reacción, los cuales conducen desde el piruvato o el fosfoenolpiruvato (productos de la glucólisis), al CO₂, CH₄ y ácidos acético, propiónico, butírico y valeriánico (AGV).
ACETATO Y CO2
El acetato y el CO₂ son producidos, a partir de los carbohidratos por numerosas especies individuales de microorganismos del rumen, mientras que otras especies los producen a partir de los ácidos propiónico, butírico y valeriánico.
Sin embargo, muchas de estas especies individuales también tienen uno o más productos finales tales como succinato, lactato, formiato, hidrógeno y etanol, que no son productos finales de la fermentación.
Estos compuestos, con la posible excepción del etanol, son metabolizados posteriormente por otras especies que dan lugar a los productos finales típicos.
AGV
Los AGV son productos finales de la fermentación microbiana y son absorbidos a través del epitelio queratinizado del rumen.
Estos ácidos son compuestos de cadena carbonada corta, que se producen durante la degradación fermentativa de los alimentos y pueden ser convertidos en glucosa, aminoácidos o ácidos grasos por las bacterias ruminales o por las células del animal.
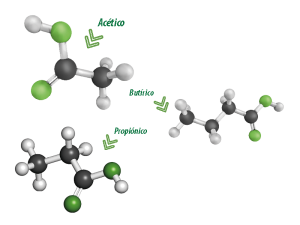

Los que más se producen en mayor cantidad son acético, propiónico y butírico, que son producidos por las enzimas intra y extracelulares de bacterias y protozoos.
La mayoría del acetato y todo el propionato son transportados al hígado, pero la mayoría del butirato se convierte en la pared del rumen a una cetona que se llama [beta]-hidroxibutirato (Figura 1).
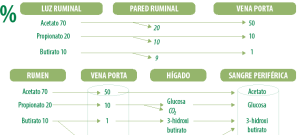



Los ácidos acético, propiónico y butírico, conforman la mayoría (>95%) de los ácidos producidos en el rumen.
También la fermentación de aminoácidos generados en el rumen produce ácidos, llamados iso-ácidos.
La energía y los iso-ácidos producidos durante la fermentación son utilizados por las bacterias para crecer (es decir, principalmente para sintetizar proteína).
![]()
El etanol, succinato, y lactato son catabolizados a productos finales: acético, propiónico y butírico por las distintas especies de microorganismos (Santini, 1994).
El acético es transportado desde el rumen por el torrente circulatorio e incorporado directamente al proto-plasma celular. Su utilización puede orientarse posteriormente ya sea hacia la obtención de energía vía acetil-CoA y ciclo de Krebs, o bien a la síntesis de grasas (tanto tejido graso como grasa en leche).
El CO₂ y CH₄ son eructados, y la energía todavía presente en el CH₄ se pierde.
Si no es necesaria para el mantenimiento de la temperatura del cuerpo, el calor producido durante la fermentación se disipa.
La degradación y fermentación de polisacáridos (hidratos de carbono complejos) en el rumen ocurre esencialmente en tres pasos:
01. Fijación
Fijación de los microorganismos a las partículas del forraje, provocando la disociación de los hidratos de carbono de la estructura de la matriz celular.
02 Hidrólisis
Hidrólisis de los polisacáridos liberados, a monosacáridos.
03 Fermentación
Fermentación intracelular de los monosacáridos, obteniéndose AGV. De la fermentación intracelular de las hexosas y pentosas se obtienen principalmente piruvato y fosfoenolpiruvato. Sobre los que actúan los microorganismos y los transforman en distintos productos finales de la fermentación ruminal a través de distintas vías metabólicas (Leng 1973).


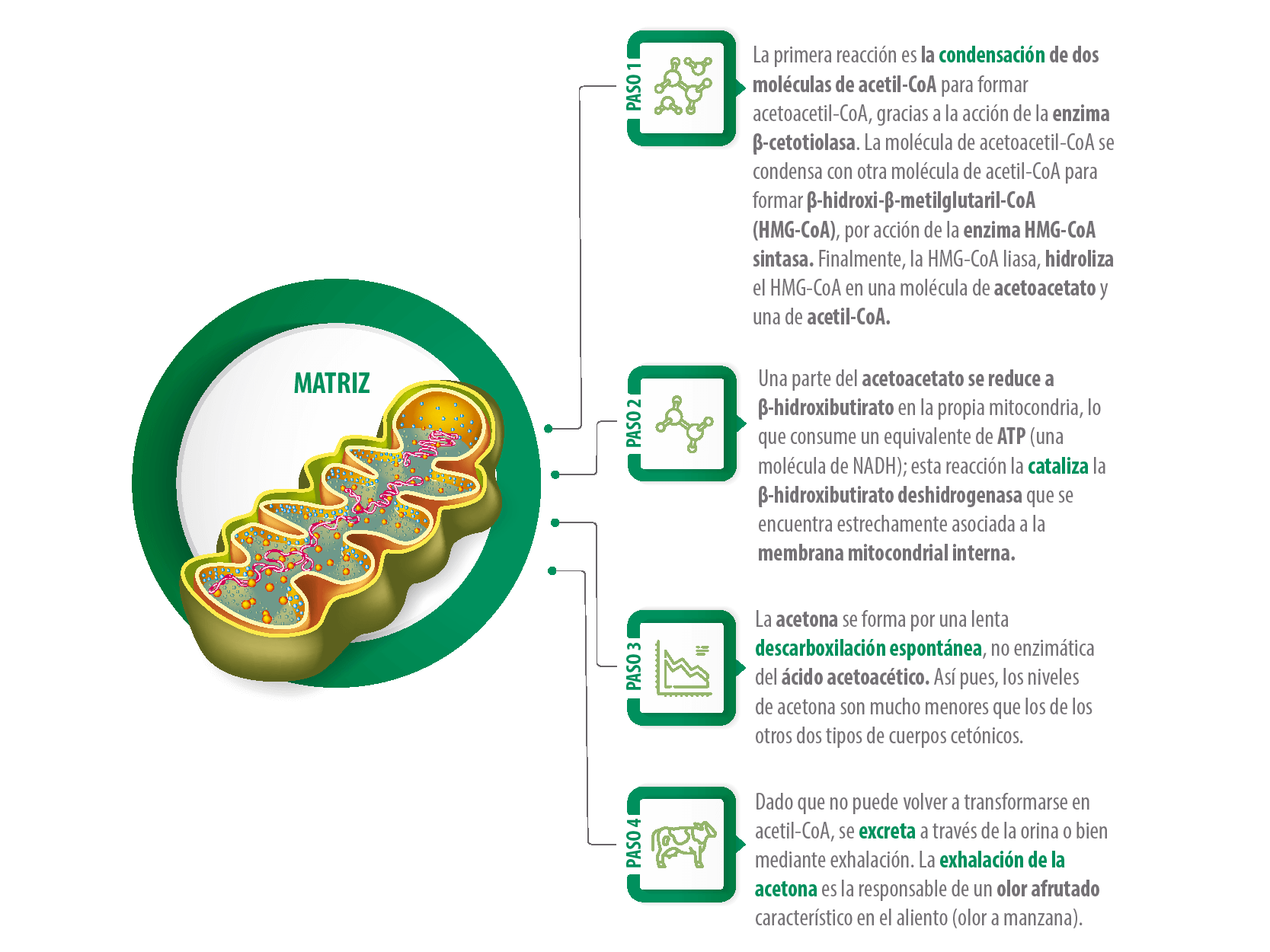
CETONAS
Las cetonas son la fuente principal de energía (combustible) para la mayoría de tejidos del cuerpo.
Las cetonas provienen principalmente del butirato producido en el rumen, pero en las etapas iniciales de lactancia vienen también de la movilización de tejidos adiposos.
El lugar primario de formación de los cuerpos cetónicos es el hígado y, en menor proporción, el riñón.
El proceso tiene lugar en la matriz mitocondrial y ocurre en diferentes pasos:

CATABOLISMO Y ANABOLISMO


Los nutrientes absorbidos en el rumen o en el intestino delgado, se dirigen principalmente al hígado y a otros órganos para su catabolismo o anabolismo, según las necesidades.
El metabolismo de los monosacáridos gira en torno al suministro y destino de las hexosas, siendo este monómero la principal fuente de energía para las células, la vía catabólica es la glucólisis, llevada a cabo en el citoplasma celular. Este proceso consta de ocho reacciones:
- La glucosa ingresa al citoplasma para ser fosforilada (adición de un grupo fosfato), a partir de ATP. Esta reacción es catalizada por la hexoquinasa. La glucosa 6-fosfato (aldohexosa) resultante abunda en todas las células, ya que la gran mayoría que ingresa al citoplasma termina siendo fosforilada, con el fin de impedir que pueda atravesar de regreso la membrana citoplasmática y difundirse al medio extracelular.
- La aldohexosa presenta isomerización y es transformada en fructosa 6-fosfato (cetohexosa). Reacción catalizada por glucosa-6-fosfato isomerasa.
- La fructosa-6-fosfato, es fosforilada a partir de ATP, en los carbonos 1 y 6 para dar lugar a la fructosa-1,6bisfosfato. Reacción catalizada por fosfofructoquinasa.
- La fructosa-1,6-bisfosfato está dividida en dos: gliceraldehido-3-fosfato y dihidroxiacetona fosfato. Reacción catalizada por aldosa.
- La triosa fosfato isomerasa cataliza la conversión de dihidroxiacetona fosfato para obtener más gliceraldehido-3-fosfato
- El gliceraldehido-3-fosfato es oxidado y fosforilado, en los carbonos 1 y 6 formando 1,3-bifosfoglicerato por gliceraldehidofosfato deshidrogenasa, posteriormente, transfiere su grupo fosfato, para sintetizar ATP y se transforma en 3-fosfoglicerato.
- El 3-fosfoglicerato presenta isomerización del C3 al C2 y es transformado en 2-fosfoglicerato por la fosfoglicerato mutasa. Posteriormente la enolasa propicia la formación de un enlace doble, eliminando una molécula de H₂O y formando fosfoenolpiruvato.
- El fosfoenolpiruvato transfiere su grupo fosfato, para sintetizar ATP y se transforma en Piruvato.


El piruvato entra en la matriz mitocondrial, por lo tanto, cada hexosa (glucosa) genera:
2 piruvatos, 2 ATPs, 2 NADH+H+ , 2 hidrogeniones y 2 moléculas de H₂O, los piruvatos se metabolizan a acetil-Coenzima A, permitiendo así su ingreso al ciclo de Krebs y produciendo 12 ATPs (Figura 2)
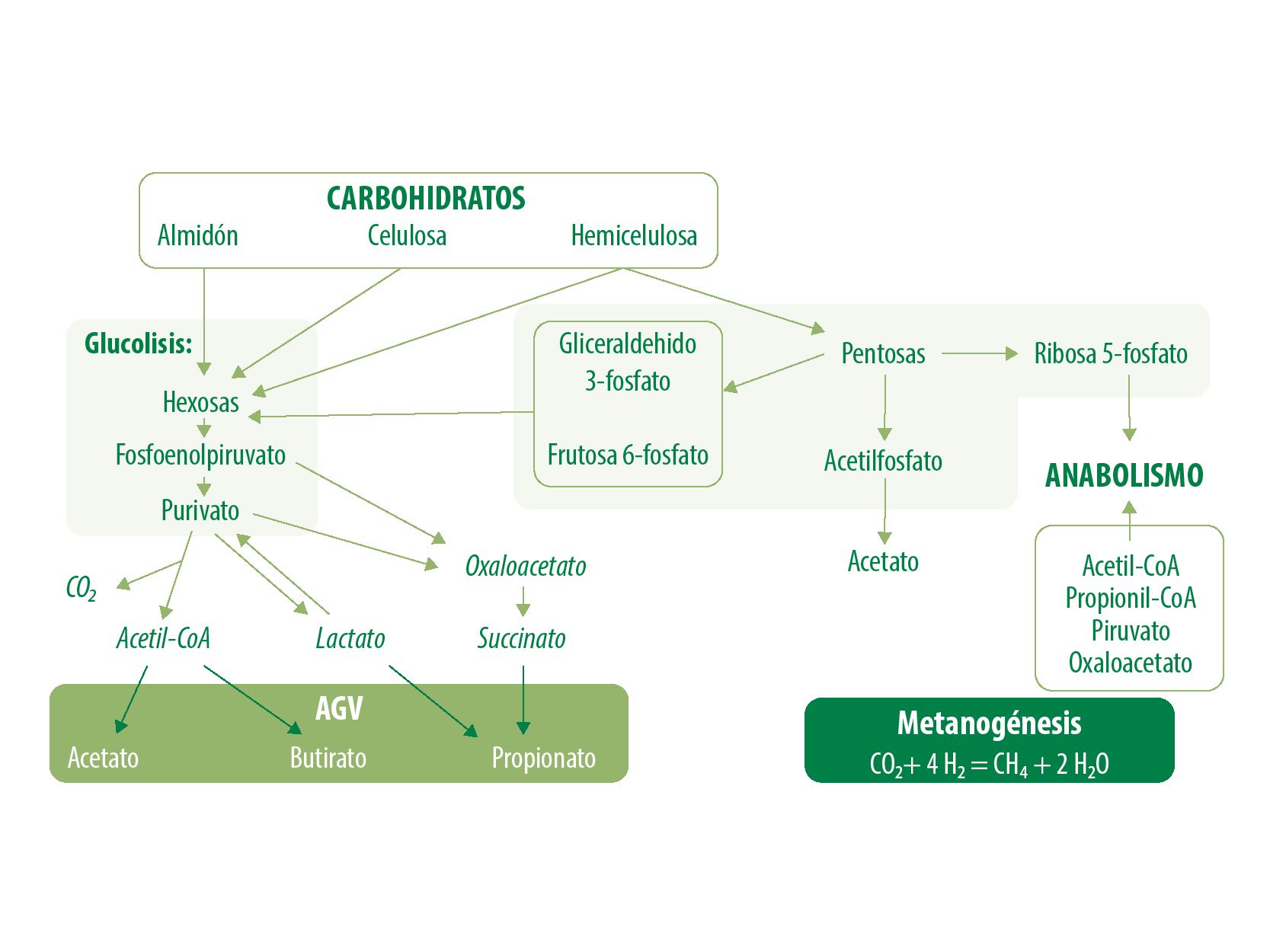
Figura 2. Metabolismo de los carbohidratos.

GLUCONEOGÉNESIS
Todo el propionato se convierte en glucosa en el hígado. Además, el hígado utiliza los aminoácidos para la síntesis de glucosa, este proceso es conocido como gluconeogénesis.
Es muy importante, porque en general no hay suficiente glucosa absorbida en el tracto digestivo y todos los azúcares encontradas en leche (lactosa aproximadamente 1575g en una vaca que produce 35 kg de leche al día) deben ser producidas por el hígado en dietas forrajeras.
La gluconeogénesis también sintetiza GLUCOSA a partir de lactato y glicerol, en el citoplasma celular y en las mitocondrias de los hepatocitos.
En esta ruta metabólica se consumen seis ATPs por cada hexosa producida y el propionato CH3-CH2-COOH es el único AGV glucogénico.


La importancia de la glucogénesis en rumiantes, se debe principalmente a que su organismo absorbe cantidades pequeñas de glucosa en el tracto digestivo y a que su capacidad de almacenar glucógeno en el hígado es más limitada que en los monogástricos.