



En esta segunda parte continuamos con el metabolismo de ácidos grasos volátiles (AGV), para terminar con los nuevos descubrimientos sobre su absorción a través del epitelio ruminal.


La digestibilidad del almidón en el total del tracto, es superior al 80%, variando de acuerdo al tipo de grano y al consumo (MS/día), la proporción que se degrada en el rumen y, la que llega intacta al duodeno.
Degradabilidad del almidón


Durante la lactancia, la glándula mamaria tiene una alta prioridad para la utilización de glucosa, aquí se utiliza principalmente para la formación de lactosa – o azúcar de la leche -, un disacárido formado por una glucosa y una galactosa (Figura 1).
La cantidad de lactosa sintetizada en la ubre, está estrechamente ligada con la cantidad de leche producida cada día.
La concentración de lactosa en la leche es relativamente constante, el agua se agrega a la cantidad de lactosa producida por las células secretoras hasta lograr una concentración de lactosa de aproximadamente 4,5%.
Así, la producción de leche en las vacas está altamente influida por la cantidad de glucosa derivada del propionato producido en el rumen.
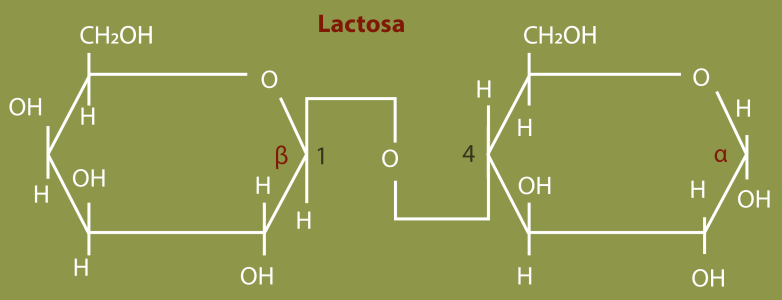

Figura 1. Lactosa
La glucosa también se convierte a glicerol que se utiliza para la síntesis de los triglicéridos de la grasa de leche. La energía requerida para la síntesis de grasa y lactosa se obtiene de la combustión de las cetonas, pero el acetato y la glucosa también pueden ser utilizadas como fuentes de combustible para las células de muchos otros tejidos.

Alrededor del 20–30% de la grasa de la leche es sintetizada en la glándula mamaria (síntesis de novo). La otra parte, que contiene parte de ácidos grasos insaturados de 16 a 22 átomos de carbono (ácidos grasos de cadena larga), viene de lípidos de la dieta y también puede proceder de la movilización de grasa corporal como sucede en el pico de la lactación.


La fuente de carbohidratos en la dieta influye en la cantidad y la relación de AGV producidos en el rumen.![]()

![]()

Metabolismo de los carbohidratos

Además, los carbohidratos no-fibrosos producen más AGV (es decir más energía) porque son fermentados más rápidamente y en forma completa.
Así, la alimentación alta en concentrados usualmente da como resultado un aumento de producción de AGV y una proporción mayor de propionato en lugar de acetato.
Cuando se alimentan los rumiantes con grandes cantidades de concentrados (o cuando se alimentan con forrajes muy molidos), el porcentaje de ácido acético se reduce debajo de 40% mientras el porcentaje de propionato aumenta a más de 40%.
En la figura 2, podemos visualizar estas diferentes vías de producción de AGV, dependiendo de la fuente de carbohidratos disponible.
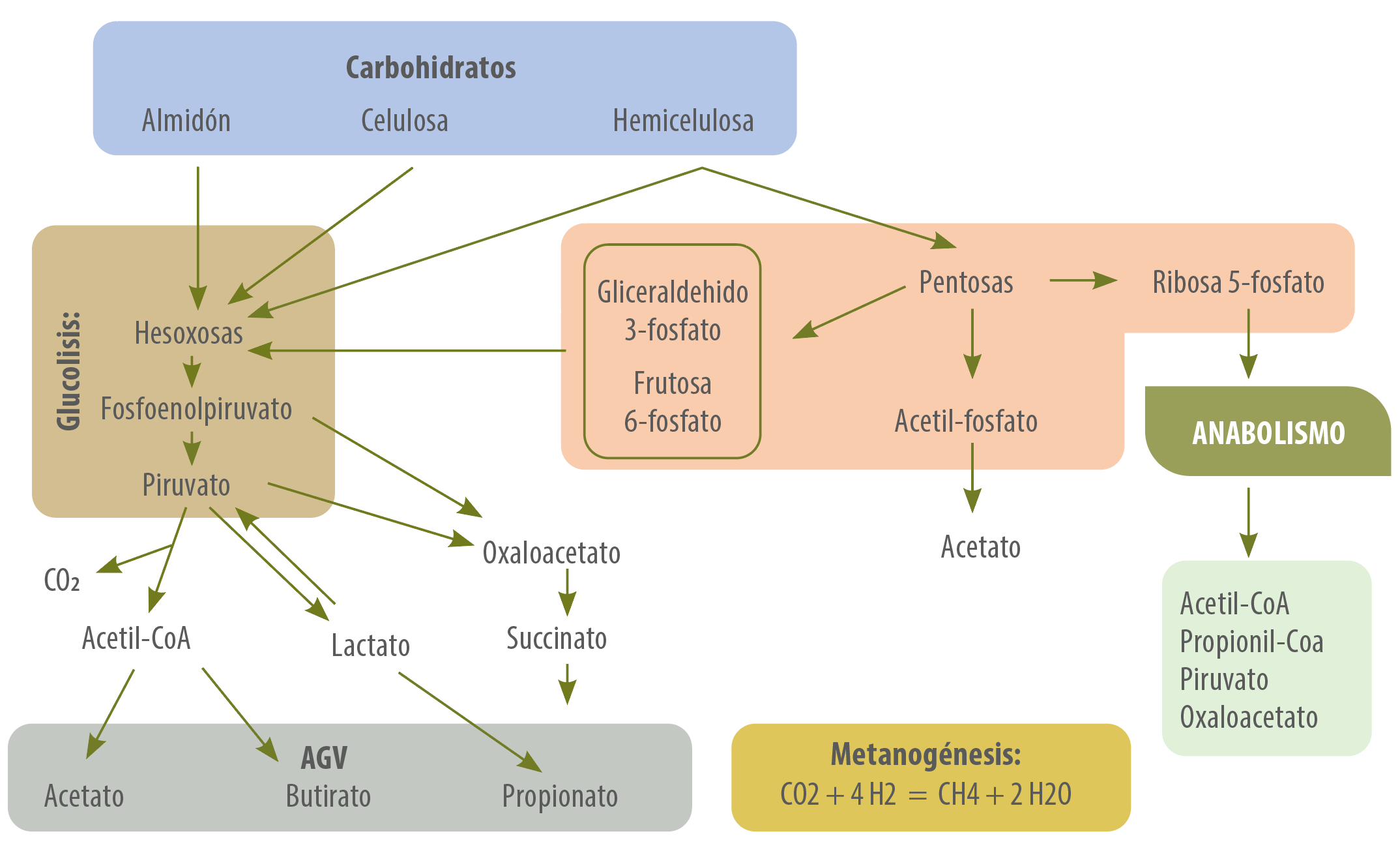

Figura 2. Metabolismo de los carbohidratos.
Esto habitualmente provoca un aumento en la producción de leche porque el suministro de glucosa proveniente de propionato se incrementa, pero el suministro de ácido acético para la síntesis de grasa puede llegar ser limitante.
En general, esta reducción en disponibilidad de ácido acético, se asocia a una reducción de producción de grasa en la leche. Además, un exceso de propionato relativo al acetato (relación acético:propiónico baja) provoca que la vaca, al utilizar la energía disponible la destine a depositar tejido adiposo, aumentando de peso corporal en lugar de sintetizar la grasa de la leche, especialmente en animales con producciones bajas.

La alimentación prolongada con este tipo de raciones puede tener un efecto negativo para la salud de la vaca, que tienden más a ser afectadas por: hígado graso, cetosis, distocias (dificultades de parición), etc. Por otro lado, niveles bajos de concentrado en la ración, en general, limita la ingestión de energía y la producción de leche.




Los AGV tienen un efecto importante en:
![]()

![]()
![]()
![]()
![]()

![]()
En cambio, el ácido propiónico es, el responsable en primer término de la síntesis de lactosa. El ácido butírico no muestra carácter específico, siendo utilizado en la síntesis de los tres principales componentes de la leche.
Para obtener en la leche un porcentaje graso fisiológicamente normal (superior al 3%) se necesita una relación acetato:pro pionato de por lo menos 3:1, y se obtiene cuando el contenido en fibra expresada en fibra neutro detergente constituye alrededor de un 20% de la materia seca de la ración.

Para engorde, la relación molar entre el acético y el propiónico es más estrecha, con una mayor concentración relativa del segundo. Esto se logra con un menor contenido de fibra, lo que permitiría la incorporación de concentrados a la ración.
![]()

La importancia del pH
En cuanto a la absorción y el metabolismo de los AGV (Figura 3), Dijkstra et al. (1993) informaron que la tasa de absorción de AGV aumentó con la disminución del pH y que el aumento en la absorción fue lineal a la reducción del pH.
Dada la falta de saturación en la tasa de absorción con el aumento de la concentración de AGV y un aumento de la absorción de AGV con pH más bajo, se pensó en gran medida que la absorción se producía a través de la difusión pasiva.
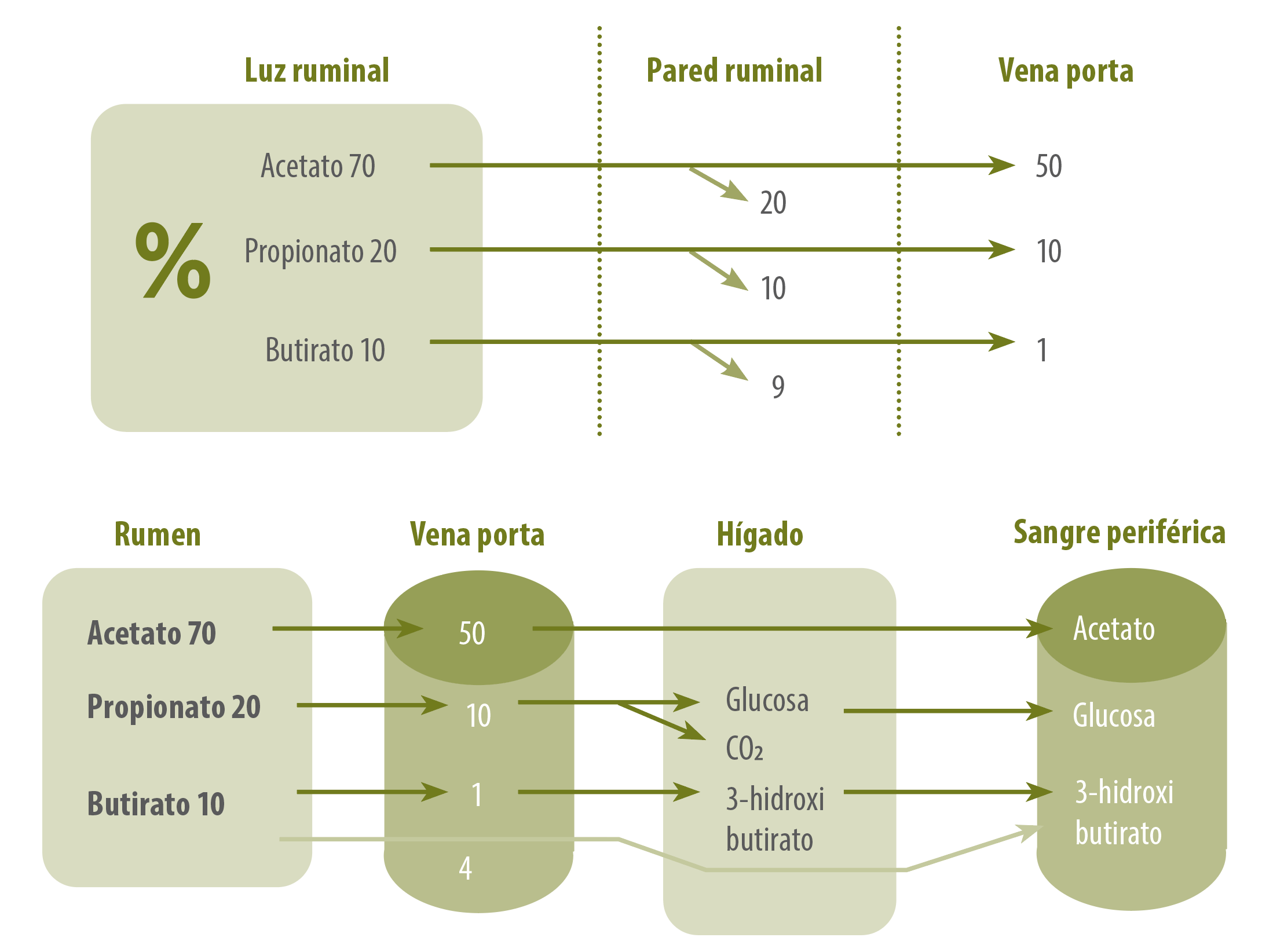

Figura 3. Absorción y metabolismo de los ácidos grasos volátiles.


Por lo tanto, una reducción en el pH aumentaría la proporción de AGV no disociado que luego podría difundirse libremente a través del epitelio ruminal.
Si bien la evidencia parecía apoyar la teoría de la difusión pasiva, existen numerosas restricciones teóricas. En primer lugar, la proporción de AGV en el estado no disociado es baja en condiciones normales de pH en el rumen.
Los ácidos grasos volátiles tienen un pKa de 4.8. Incluso con valores de pH de 5,8, más del 90% del AGV estaría en estado disociado y, por lo tanto, solo una pequeña fracción sería lipofílica y podría esperarse que la tasa de absorción de los AGV avanzara lentamente.
Aunque la proporción relativa de AGV no disociado sería baja, se sugirió que había una micro zona de pH ácido al lado luminal de los epitelios ruminales. Esta zona podría favorecer que AGV esté en el estado no disociado adyacente al epitelio, promoviendo así la absorción.


Sin embargo, se ha encontrado que el pH en la superficie de los epitelios es básico con valores que oscilan entre 7.47 y 7.68 dependiendo de las condiciones de incubación.
Las limitaciones al modelo de difusión pasiva exclusiva también se extienden a las diferencias para la lipofilia de los AGV individuales.
![]()




Los mecanismos predominantes incluyen:
1) AGV- / HCO3 – intercambio aniónico,
2) difusión pasiva,
3) absorción de AGV sensible al nitrato,
4) transporte de AGV acoplado a protones y,
5) transporte electrogénico de AGV
Si bien estos son los principales mecanismos de absorción, se requieren otros procesos como el intercambio de Na/H y la importación de bicarbonato en la célula para permitir el mantenimiento del pH intracelular y promover la absorción de AGV.
![]()
Sin embargo, sobre la base de los datos disponibles, parece que para el acetato la proporción representada por el transporte dependiente del bicarbonato, el sensible a los nitratos y la difusión pasiva es del 42 al 57%, del 0 al 14% y del 29 al 59%, respectivamente.
Para el butirato, la proporción registrada por el transporte dependiente del bicarbonato, el sensible a los nitratos y la difusión pasiva son de 24 a 46%, de 0 a 4% y de 25 a 76%, respectivamente.
En el rumen, la gran mayoría del AGV estará en estado disociado (AGV-). La absorción de AGV- ocurre a cambio de HCO3- en un proceso electro-neutro que está mediado por una serie de intercambiadores de aniones potenciales.
Este mecanismo proporciona una fuente de bicarbonato al ambiente ruminal donde puede neutralizar un protón a través de la reacción de anhidrasa carbónica que produce dióxido de carbono y agua.
Modelo de absorción de AGV


Modelo que resume el conocimiento actual de la absorción de AGV en relación con la estabilización del pH ruminal.
1 La absorción por difusión del AGV facilita la eliminación de un protón asociado con el AGV. Este protón se disociará rápidamente en el citosol donde puede ser exportado por intercambios de sodio/hidrógeno
7 8 o acoplado con metabolitos de AGV (por ejemplo, cuerpos cetónicos y lactato) a través del transportador de monocarboxilato
4 El AGV disociado puede absorberse en un mecanismo de intercambio aniónico, proporcionando así una fuente de bicarbonato al contenido ruminal
2 Este bicarbonato puede neutralizar un protón a través de la reacción de la anhidrasa carbónica, estabilizando así el pH ruminal. El suministro de bicarbonato a los epitelios viene de la sangre
5 6 El AGV también se puede absorber a través de una vía sensible al nitrato
3 y se puede exportar a la sangre a través de un canal dependiente de gasto energético
10 Tenga en cuenta que el modelo no muestra la complejidad estructural de los epitelios ruminales, incluido el número de estratos y células dentro de los estratos.
https://animal.ifas.ufl.edu/apps/dairymedia/rns/2014/penner.pdf
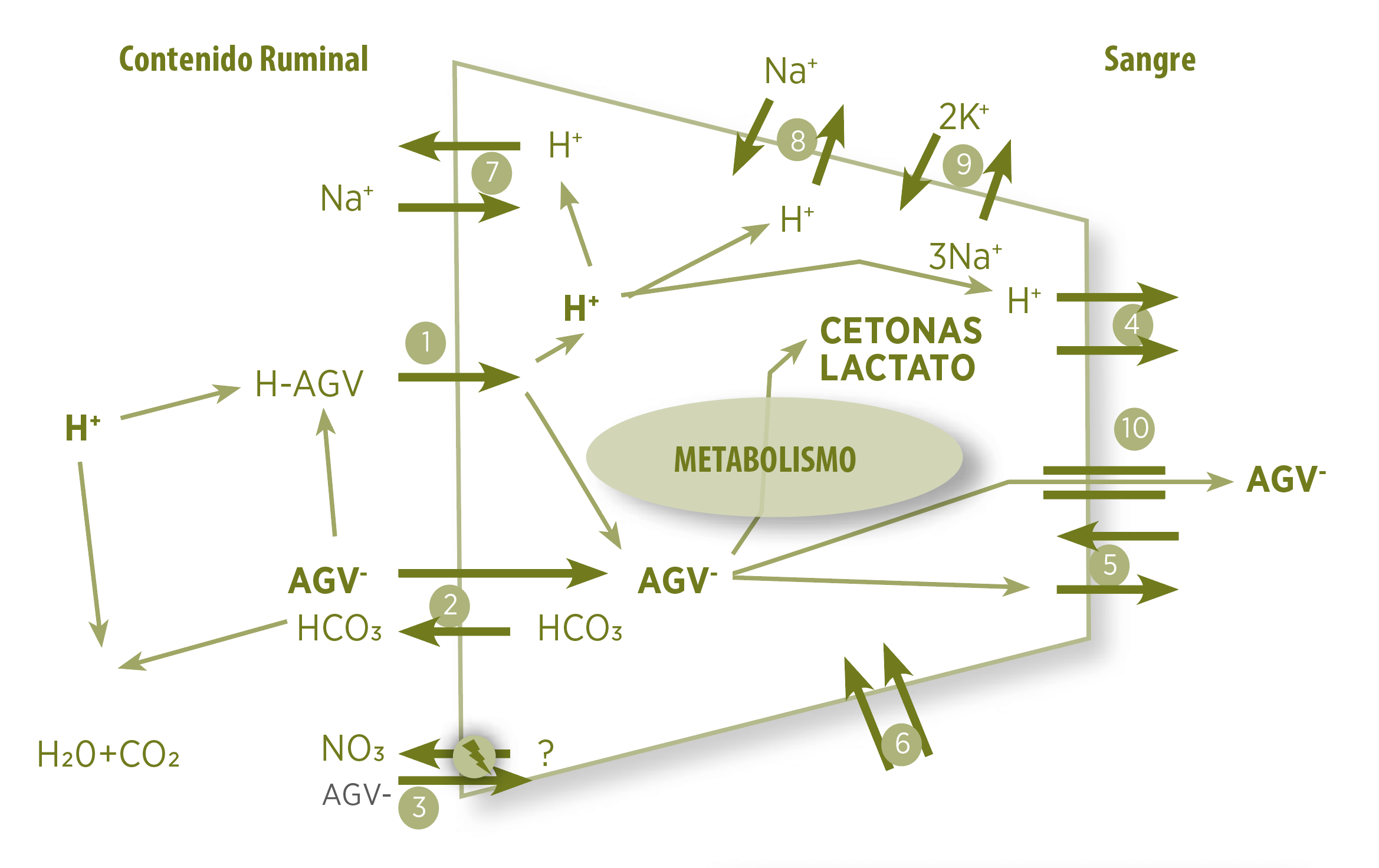

Figura 4. Modelo de absorción de ácidos grasos volátiles.