La cría y recría de terneras lecheras es un proceso clave para asegurar el futuro productivo del rebaño. El objetivo principal es lograr un adecuado desarrollo corporal y del tejido mamario, lo que permitirá a las terneras expresar su potencial de producción en la edad adulta. Este proceso se divide en varias etapas que se esquematizan en la Figura 1.

Figura 1. Esquema de las diferentes etapas de la cría y recría de terneras lecheras donde se representa el nacimiento (momento en que el calostrado es fundamental), el desleche (aproximadamente a los 2 meses de edad), la pubertad (a la que se llega a los 10 meses aproximadamente, cuando la ternera alcanza el 40-45% de su peso vivo adulto (PVa)), el momento ideal para el primer servicio (alrededor de los 15 meses, cuando la ternera alcanza el 55% de su PVa). Todo este proceso permitirá llegar a los 2 años al primer parto con un peso óptimo (85% del PVa).
Digestión en las primeras etapas de la vida
Las crías de rumiantes nacen con preestómagos de tamaño reducido y carecen de microorganismos suficientes capaces de degradar fibras vegetales, lo que les impide obtener energía a través de la fermentación microbiana en el tracto digestivo.
Por esta razón, durante las primeras 3 a 4 semanas de vida, los procesos digestivos de los pre-rumiantes son similares a los de un monogástrico, dependiendo principalmente de enzimas digestivas propias que actúan en el abomaso y el intestino para procesar los nutrientes.
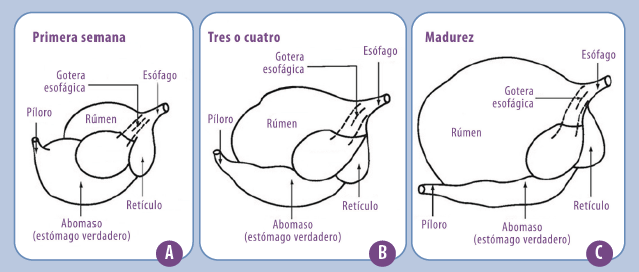
Figura 2. Desarrollo esquematizado del aparato digestivo del rumiante en la primera semana de vida (A), a los 3-4 meses (B) y en la madurez (C). En la primera semana de vida, el rumen representa el 25% de la capacidad total del tracto digestivo, mientras que en la madurez llega al 80%. Lo contrario sucede con el abomaso, que en la primera semana de vida representa el 60% de la capacidad digestiva total, y en el bovino adulto no superará el 8% (extraído de Jones y Heinrichs, https://extension.psu.edu/feeding-the-newborn-dairy-calf#section-27)
En esta etapa, las enzimas digestivas son limitadas en cantidad y variedad, pero son suficientes para digerir los componentes principales del calostro y la leche, como la caseína, lactoglobulinas, triglicéridos y lactosa, que constituyen los alimentos naturales de las crías durante sus primeras semanas de vida. A medida que la dieta se diversifica, el espectro de enzimas digestivas se amplía, adaptándose a los nuevos sustratos.
|
|
![]() El tracto digestivo de los rumiantes tiene una disponibilidad muy restringida de enzimas hasta aproximadamente los 21 días de edad. El complejo enzimático del recién nacido está compuesto por una enzima proteolítica con actividad (la quimosina), lipasas pregástricas y pancreáticas (aunque esta última tiene una actividad débil al principio), y la lactasa, que es la enzima glicolítica encargada de digerir la lactosa, el carbohidrato principal de la leche.
El tracto digestivo de los rumiantes tiene una disponibilidad muy restringida de enzimas hasta aproximadamente los 21 días de edad. El complejo enzimático del recién nacido está compuesto por una enzima proteolítica con actividad (la quimosina), lipasas pregástricas y pancreáticas (aunque esta última tiene una actividad débil al principio), y la lactasa, que es la enzima glicolítica encargada de digerir la lactosa, el carbohidrato principal de la leche.
![]() En esta etapa, las maltasas (que digieren carbohidratos de cadena corta) son incipientes, y las amilasas (que permiten digerir el almidón) aún no están presentes (Figura 3). Con este paquete enzimático inicial, los rumiantes están capacitados para digerir los nutrientes de la leche, pero no mucho más.
En esta etapa, las maltasas (que digieren carbohidratos de cadena corta) son incipientes, y las amilasas (que permiten digerir el almidón) aún no están presentes (Figura 3). Con este paquete enzimático inicial, los rumiantes están capacitados para digerir los nutrientes de la leche, pero no mucho más.
A lo largo de los primeros días, los rumiantes desarrollan gradualmente su batería enzimática, volviéndose capaces de digerir adecuadamente proteínas y lípidos. Sin embargo, hasta aproximadamente las 3 semanas de vida, la secreción de enzimas capaces de degradar almidones y otros hidratos de carbono distintos a la lactosa es escasa.
Por ello, es fundamental que, al emplear sustitutos lácteos desde el inicio de la vida (después del descalostrado), estos cumplan con las siguientes condiciones:
![]() La fuente de proteína sea origen lácteo o de plasma animal y si se utilizan fuentes vegetales deben de ser correctamente hidrolizadas e incluidas en bajas proporciones.
La fuente de proteína sea origen lácteo o de plasma animal y si se utilizan fuentes vegetales deben de ser correctamente hidrolizadas e incluidas en bajas proporciones.
![]() No contengan almidón. La principal fuente de glúcidos es la lactosa que normalmente forma parte del sustituto en un 45%.
No contengan almidón. La principal fuente de glúcidos es la lactosa que normalmente forma parte del sustituto en un 45%.
![]() Los lípidos, grasas o aceites deben de ser emulsificados para que puedan estar en suspensión en el líquido.
Los lípidos, grasas o aceites deben de ser emulsificados para que puedan estar en suspensión en el líquido.
![]() Los insumos provengan principalmente de la leche o hayan recibido un tratamiento específico que garantice una digestión correcta por parte de las terneras.
Los insumos provengan principalmente de la leche o hayan recibido un tratamiento específico que garantice una digestión correcta por parte de las terneras.
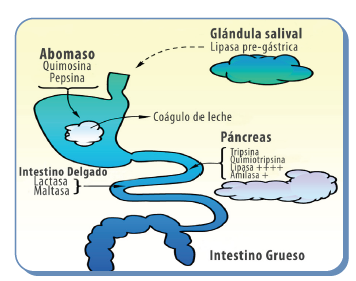
Figura 3. Enzimas digestivas segregadas por el pre-rumiante al lumen del tracto digestivo (extraído de Garin et al., 2003)
Un adecuado calostrado es clave
El calostro es la primera fuente de nutrientes y anticuerpos para las terneras. Es fundamental que las terneras ingieran calostro dentro de las primeras 24 horas de vida. Recordemos que el tipo de placenta que presentan los bovinos (epiteliocorial), impide el paso de macromoléculas, por lo que es una especie prácticamente agammaglobulinémica al nacimiento, necesitando la ingestión y absorción de calostro rico en anticuerpos y otros factores inmunes que aporten una inmunidad pasiva.
Se recomienda que las terneras consuman entre 150-200 g de IgG (inmunoglobulina G) en este período (NASEM, 2021). Para ello, tengamos en cuenta que no todos los calostros son iguales, y que la concentración de IgG puede variar mucho.
| En un estudio realizado en Estados Unidos, Heinrichs et al. (2020) observaron una enorme variación entre calostros y contaminación por E. Coli en más del 50% de los calostros analizados. Es por ello por lo que, en general se observa que tras la pasteurización mejora la absorción de Ig. En realidad, el tratamiento térmico no provoca la mejora per se, pero elimina la carga microbiana que interfiere con la absorción de las Ig. |
Por otra parte, estudios han demostrado que el calostro no solo proporciona inmunidad, sino que también contiene muchas y diversas proteínas (más de 62), es rico en energía, péptidos bioactivos, hormonas y factores de crecimiento (relaxina, insulina, IGF1, IGF2, etc.) que favorecen el desarrollo digestivo y general de la ternera (Kargar et al., 2020).
| Por ello, el suministro de calostro hasta las 72 horas de nacida la ternera favorece el crecimiento y la maduración de las vellosidades intestinales, lo cual va a resultar muy útil en el momento de la transición, cuando se vea desafiada en su pasaje de pre-rumiante a rumiante. |
Más aún, después del calostro, actualmente se enfatiza en la recomendación de suministrar “leche de transición” durante los primeros 3-4 días de vida. Esta leche que es la producida en los días posteriores al calostro, favorece el desarrollo intestinal, aumenta el tamaño de las vellosidades y mejora la salud general de las terneras (Van Soest et al., 2022).
¿Por qué importa el crecimiento?
En todos los animales (incluídos los humanos) es sabido que el crecimiento en las primeras etapas de vida es fundamental para el desempeño futuro, y las terneras lecheras no son la excepción.
 En las terneras, un mayor crecimiento en las primeras semanas de vida está asociado con un mayor desarrollo del tejido mamario y una mayor producción de leche en la primera lactancia (Brown et al., 2005). Parecería ser que estos efectos positivos en la producción de leche se observan a partir de ganancias de peso de 0,5 kg/día (Gelsinger et al., 2016).
En las terneras, un mayor crecimiento en las primeras semanas de vida está asociado con un mayor desarrollo del tejido mamario y una mayor producción de leche en la primera lactancia (Brown et al., 2005). Parecería ser que estos efectos positivos en la producción de leche se observan a partir de ganancias de peso de 0,5 kg/día (Gelsinger et al., 2016).
A partir del concepto de que la nutrición en las primeras etapas es clave y de que tradicionalmente, las terneras lecheras que se crían separadas de su madre ingieren menos leche de la que tomarían si estuvieran al pie de la madre, ha surgido, hace ya algunos años, el concepto de cría acelerada de terneras.
 Básicamente, la cría acelerada intenta duplicar el peso de las terneras a los 60 días de vida, suministrando más alimento lácteo (hasta el doble) en comparación con la cría tradicional (Drackley, 2008; Kertz et al., 2017). Este enfoque ha demostrado mejorar el crecimiento y el desarrollo de las terneras, aunque también puede reducir el consumo de concentrado durante la etapa de lactancia (Gelsinger et al., 2016). Básicamente, la cría acelerada intenta duplicar el peso de las terneras a los 60 días de vida, suministrando más alimento lácteo (hasta el doble) en comparación con la cría tradicional (Drackley, 2008; Kertz et al., 2017). Este enfoque ha demostrado mejorar el crecimiento y el desarrollo de las terneras, aunque también puede reducir el consumo de concentrado durante la etapa de lactancia (Gelsinger et al., 2016). |
Del tema cría acelerada de terneras, sus posibles ventajas e inconvenientes, nos ocuparemos en la parte II de esta entrega. Ahora nos centraremos en aspectos generales de cómo impacta la dieta y el crecimiento en el desarrollo de las terneras como futuras vacas lecheras.
Hay evidencias que permiten suponer, sin embargo, que la acumulación de tejido adiposo en torno a la glándula mamaria estaría asociada a la relación de energía y proteína de la dieta más que a la densidad energética o las ganancias diarias de PV.
Es muy importante, cuando se trata de terneras que esperamos tengan el óptimo desarrollo mamario para producir con todo su potencial cuando adultas, que la dieta no sea deficitaria en proteína. Recordemos que una ternera lechera no debe “engordar”, sino desarrollarse adecuadamente y debe crecer en altura.
| Es importante entonces que no falte la proteína en la dieta y que la energía no sobre. El nuevo NASEM (2021) propone que la relación entre proteína metabolizable (PM) y la energía metabolizable (EM) de la dieta dependerá del peso vivo de la ternera y de su relación con el peso adulto. Así, vacas de biotipos grandes y, de menos edad, necesitarán más gramos de PM por Mcal de EM (figura 4). |

Figura 4. Relación proteína metabolizable (PM) y energía metabolizable (EM) necesaria para el adecuado crecimiento de terneras lecheras según NASEM (2021). Las diferentes curvas corresponden a diferentes pesos de vaca adulta (500, 600, 700 u 800 kg de peso vivo (PV) adulto). Nótese que cuanto más grande el peso adulto estimado y más joven (con menor peso) sea la ternera, la dieta deberá tener una mayor proporción de proteína en la dieta.
También es importante apelar al conocimiento básico cuando de alimentar nuestras terneras se trata. Y en este caso podemos recurrir a la fisiología del crecimiento de los animales.
![]() Todos los animales pasan por etapas diferentes durante el crecimiento. Esquemáticamente, se denomina etapa de crecimiento “isométrico” a un crecimiento en el que todas las partes del cuerpo crecen a la misma tasa proporcional. Es decir, no hay cambios en las proporciones corporales.
Todos los animales pasan por etapas diferentes durante el crecimiento. Esquemáticamente, se denomina etapa de crecimiento “isométrico” a un crecimiento en el que todas las partes del cuerpo crecen a la misma tasa proporcional. Es decir, no hay cambios en las proporciones corporales.
![]() Mientras tanto, crecimiento “alométrico” se refiere a un crecimiento en el que diferentes partes del cuerpo crecen a tasas diferentes, lo que resulta en cambios en las proporciones corporales.
Mientras tanto, crecimiento “alométrico” se refiere a un crecimiento en el que diferentes partes del cuerpo crecen a tasas diferentes, lo que resulta en cambios en las proporciones corporales.
En el caso de las terneras lecheras, hace ya algunas décadas se describió que el crecimiento de la glándula mamaria de las vaquillonas desde el nacimiento hasta el primer parto se puede dividir en cuatro etapas:
![]() Una etapa inicial de crecimiento isométrico entre el nacimiento y los 2-3 meses de vida.
Una etapa inicial de crecimiento isométrico entre el nacimiento y los 2-3 meses de vida.
![]() Una fase de crecimiento alométrico que se extiende hasta la pubertad (desde el destete a los 9-10 meses).
Una fase de crecimiento alométrico que se extiende hasta la pubertad (desde el destete a los 9-10 meses).
![]() Una nueva fase isométrica hasta el último tercio de gestación (19-20 meses de vida)
Una nueva fase isométrica hasta el último tercio de gestación (19-20 meses de vida)
![]() Una nueva fase de crecimiento alométrico desde los 20 meses al parto (Sinha y Tucker, 1969).
Una nueva fase de crecimiento alométrico desde los 20 meses al parto (Sinha y Tucker, 1969).
| Durante las etapas de crecimiento alométrico, especialmente entre el destete y la pubertad, el tejido mamario está en plena “expansión”. Se está desarrollando el parénquima mamario (tejido secretor de leche) y este es muy sensible a la dieta. Además, en la etapa prepuberal, hormonas como el IGF-1 (factor de crecimiento insulínico tipo 1) y los estrógenos juegan un papel clave en el desarrollo mamario. |
Estas hormonas están influenciadas por la nutrición, especialmente por la ingesta de proteína y energía. Es por ello que, en esta etapa resulta especialmente importante cuidar la relación proteína/energía en la dieta.
Si esta se desbalancea en favor de la energía (o le falta proteína a la dieta) corremos el riesgo de que la ubre se engrase y la proporción de tejido secretor sea menor.
La transición de pre-rumiante a rumiante: momento clave en la vida de la ternera
![]() Uno de los objetivos fundamentales en la cría de las terneras es favorecer la transición de lactante a rumiante. Para ello, el consumo de sólidos progresivo es fundamental. Sin embargo, el concepto sobre el sólido que acompaña la leche está cambiando.
Uno de los objetivos fundamentales en la cría de las terneras es favorecer la transición de lactante a rumiante. Para ello, el consumo de sólidos progresivo es fundamental. Sin embargo, el concepto sobre el sólido que acompaña la leche está cambiando.
![]() Históricamente, el alimento sólido se formulaba solamente con el objetivo de sustituir a los lácteos (alimentos caros) y así poder retirarlos rápido, para suministrar alimentos más baratos. Con el nuevo paradigma de alimentar lo mejor posible al rumiante para optimizar el crecimiento y que tenga una mejor vida productiva, el reto de formular un concentrado que favorezca la transición se vuelve más importante.
Históricamente, el alimento sólido se formulaba solamente con el objetivo de sustituir a los lácteos (alimentos caros) y así poder retirarlos rápido, para suministrar alimentos más baratos. Con el nuevo paradigma de alimentar lo mejor posible al rumiante para optimizar el crecimiento y que tenga una mejor vida productiva, el reto de formular un concentrado que favorezca la transición se vuelve más importante.
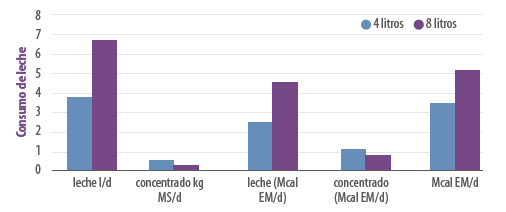
Figura 5. Consumo de leche y concentrado en terneras Holstein bajo un sistema de cría tradicional (4 litros, barras azules) y de cría acelerada (8 litros, barras rojas) (De Trinidad et al., 2014). Obsérvese que las terneras de cría tradicional, al consumir menos leche consumen más concentrado, y que, por lo tanto, la ingesta de energía proveniente del concentrado es mayor en las terneras criadas bajo un sistema tradicional. Sin embargo, cuando se calcula la ingesta energética total, resulta que la leche siempre representa la mayor ingesta de energía. En este trabajo, la leche representó el 86% de la energía metabolizable (EM) ingerida en las terneras criadas en forma acelerada, mientras que en las terneras de cría tradicional representó el 73% de la EM ingerida. En ambos casos, la mayor ingesta de energía fue a través de la leche.
Debemos tener en cuenta que, durante la etapa de lactante, sea cual sea el sistema de cría que sigamos (acelerada o tradicional), la leche o sustituto representa el principal aportador de energía con que cuentan las terneras. Esto se ejemplifica en la Figura 5 proveniente de un trabajo en el que se compara la cría tradicional con la cría acelerada.
Con este panorama, queda claro que debemos promover el desarrollo de un rumen funcional en la futura vaca ¿Qué implica esto? Podemos resumirlo en 3 puntos:
![]() Promover el desarrollo del tamaño del órgano
Promover el desarrollo del tamaño del órgano
![]() Promover el desarrollo papilar
Promover el desarrollo papilar
![]() Promover el desarrollo de una microbiota diversa y activa, capaz de fermentar carbohidratos complejos.
Promover el desarrollo de una microbiota diversa y activa, capaz de fermentar carbohidratos complejos.
Sabemos que la ingesta de concentrado de iniciación es imprescindible para el desarrollo de un rumen funcional, y que algunos productos de fermentación ruminal, como el butirato, son principales responsables del desarrollo del epitelio ruminal.
Sin embargo, como veremos en la próxima entrega, no debemos olvidar el forraje. Hay estudios que han demostrado que las terneras que consumen forraje tienen un ambiente ruminal más favorable, con mayor diversidad microbiana y un pH más cercano al neutro.
Además, el forraje grosero promueve el consumo de concentrado y mejora el bienestar de las terneras (Castells et al., 2013, Antúnez et al., 2024).
En definitiva
La inversión en la cría y recría de terneras lecheras es fundamental para asegurar el éxito futuro del rebaño.
Un manejo nutricional adecuado en las primeras etapas de vida, incluyendo un buen calostrado, una transición efectiva a rumiante y un crecimiento óptimo, permitirá a las terneras expresar su máximo potencial productivo en la edad adulta.
Bibliografía
Antúnez-Tort G., Bach A., Cajarville C., Vidal M., Fàbregas F., Ahangarani M.A., Terré M. 2023. Benefits of barley straw as a forage for dairy calves before and after weaning. J. Dairy Sci. 106: 7578-7590. https://doi.org/10.3168/jds.2023-23401.
Brown E. G., VandeHaar M. J., Daniels K. M., Liesman J. S., Chapin L. T., Forrest J. W., Akers R. M., Pearson R. E., Nielsen M. W. 2005. Effect of increasing energy and protein intake on mammary development in heifer calves. J. Dairy Sci. 88: 595–603. https://doi: 10.3168/jds.S0022-0302(05)72723-5.
Castells L., Bach A., Aris A., Terré M.. 2013 Effects of forage provision to young calves on rumen fermentation and development of the gastrointestinal tract. J. Dairy Sci., 96: 5226-5236. https://doi.org/10.3168/jds.2012-6419.
De Trinidad S., Cajarville C. Garofalo E. G., Dayuto J. Martinez A., Pereira J., Morales T., Pla M. Priore L., Mendoza A. 2014. Efecto del aumento del nivel de oferta de leche sobre el consumo de energía metabolizable en terneras Holstein. En Proceedings de XLII Jornadas Uruguayas de Buiatría. https://bibliotecadigital.fvet.edu.uy/bitstream/handle/123456789/1960/JB2014_231-233.pdf?sequence=1&isAllowed=y.
Drackley J. K. 2008. Calf Nutrition from Birth to Breeding. Veterinary Clinics of North America: Food Animal Practice, 24:55-86. https://doi.org/10.1016/j.cvfa.2008.01.001.
Garín D., Cajarville C., Corso C., Prieto M. 2003. Digestion y alimentacion en los pre-rumiantes (Cap 3), en: Curso a distancia sobre nutricion de rumiantes. Modulo 5. Facultad de Veterinaria, UdelaR, Uruguay.
Gelsinger S. L., Heinrichs A. J., Jones C. M. 2016. J. Dairy Sci. 99: 6206-6214. https://doi.org/10.3168/jds.2015-10744.
Guilloteau P., Le Huërou-Luron I., Malbert C.H., Toullec R. 1995. Les sécrétions digestives et leur régulation. En: R. Jarrige et al (eds.). Nutrition des ruminants domestiques. Ingestion et digestión. INRA Editions, Paris.
Heinrichs A. J., Jones C. M., Erickson P. S., Chester-Jones H., Anderson J. L. 2020. J. Dairy Sci. 103:5694–5699. https://doi.org/10.3168/jds.2019-17408
Jones C.M., Heinrichs J. 2025. Feeding the Newborn Dairy Calf. Penn State Extension. Disponible en: https://extension.psu.edu/feeding-the-newborn-dairy-calf#section-27 (acceso 05/03/2025)
Kargar S., Roshan M., Ghoreishi S., Akhlaghi A., Kanani M., Abedi Shams-Abadi A. R., Ghaffari M. H. 2020. J. Dairy Sci. 103:8130–8142. https://doi.org/10.3168/jds.2020-18355.
Kertz, A. F., T. M. Hill, J. D. Quigley, A. J. Heinrichs, J. G. Linn, and J. K. Drackley. 2017. A 100-Year Review: Calf nutrition and management. J Dairy Sci. 100:10151–10172. http://doi:10.3168/jds.2017-13062
NASEM, National Academies of Sciences, Engineering, and Medicine. 2021. Nutrient requirements of dairy cattle: Eighth revised edition. Washington, DC: The National Academies Press. https://doi.org/10.17226/25806.
Sinha Y. N., Tucker H. A. 1969. Mammary Development and Pituitary Prolactin Level of Heifers from Birth through Puberty and during the Estrous Cycle. J. Dairy Sci.52: 507-512. https://doi.org/10.3168/jds.S0022-0302(69)86595-1.
Van Soest B, Weber Nielsen M, Moeser AJ, Abuelo A, VandeHaar MJ. 2022. Transition milk stimulates intestinal development of neonatal Holstein calves. J. Dairy Sci. 105:7011-7022. https://doi.org/10.3168/jds.2021-21723.