Para que o intestino apresente uma boa funcionalidade, além da integridade intestinal, é necessária uma microbiota em equilíbrio, que é formada por micro-organismos não patogênicos e de fundamental importância para o metabolismo e absorção de nutrientes e outros compostos consumidos pelos animais. Uma microbiota saudável apresenta alta diversidade de gêneros microbianos em perfeito equilíbrio, permitindo um aumento da capacidade metabólica do intestino. A microbiota benéfica auxilia a digestão e absorção dos nutrientes, produz vitaminas que são utilizadas pelo hospedeiro e diminui, por competição exclusiva e/ou liberação de substâncias, a proliferação de agentes patogênicos. Esta microbiota nativa produz ácidos graxos de cadeia curta e ácido lático. Esses ácidos orgânicos diminuem o pH da excreta, favorecendo a inibição das bactérias patogênicas e estimulando a proliferação dos enterócitos. Com isso pode ocorrer uma melhoria nas estruturas e integridade das células, aumentando a capacidade de absorção dos nutrientes e consequente melhora no desempenho. Já as bactérias patogênicas causam inflamação na mucosa intestinal, geram metabólitos tóxicos, e propiciam o aparecimento de enfermidades, como é o caso da Salmonella, E. coli, Clostridium, Staphylococcus, Pseudomonas.
A microbiota intestinal dos animais tem ainda um importante papel na regulação da resposta do sistema imune, pois, além de modular vários processos fisiológicos, nutrição, metabolismo e exclusão de patógenos, pode alterar a fisiopatologia de doenças conferindo resistência ou promover infecções parasitárias entéricas. As bactérias naturais do intestino atuam como adjuvantes moleculares que fornecem imunoestimulação indireta ajudando o organismo a se defender contra infecções.
Diversos aditivos tem a capacidade de modular positivamente ou negativamente a funcionalidade intestinal e, em consequência, influenciar a saúde e desempenho animal. Alguns atuam como moduladores da microbiota, do sistema imunológico e da integridade intestinal, sendo chamados compostos nutracêuticos, que incluem prebióticos, probióticos, ácidos orgânicos, simbióticos, enzimas exógenas, ácidos graxos poli-insaturados e fitobióticos (Huyghebaert et al., 2011). Dentre eles estaca-se a levedura que através de seus compostos altera a constituição da microbiota por diversos mecanismos, modula a resposta imune, a produção de enzimas, vitaminas e outros metabólitos que afetam as bactérias intestinais quando usados como prebióticos. A parede celular de levedura Saccharomyces cerevisiae (ImmunoWall®®, ICC Brazil) oriunda do processo de fermentação da cana-de-açúcar para produção de etanol, contém em torno de 35% de β-glucanas (1,3 e 1,6), e 20% de mananoligossacarídeos (MOS). As β-glucanas são reconhecidas pelas células fagocíticas (Petravić-Tominac et al., 2010), estimulando-as a produzir citocinas que iniciarão uma reação em cadeia para induzir uma imunomodulação e melhorar a capacidade de resposta do sistema imunológico inato. Já o MOS, possui uma capacidade de aglutinação de patógenos que possuem fímbrias, tais como diversas cepas de Salmonellae Escherichia coli. Um recente estudo de Beirão et al.(2018) onde frangos de corte foram alimentados com dietas suplementadas com ImmunoWall®® (0,5 kg/ton) e infectados aos dois dias de idade com SalmonellaEnteritidis [SE] (via oral na dosagem de 108UFC/ave), mostrou alterações na microbiota e histopatologia do íleo e ceco.
Os tratamentos utilizados no estudo foram: controle (sem aditivos e sem desafio); desafio com Salmonella(108UFC/ave); ImmunoWall® 0,5 kg/ton(sem desafio) desafio com Salmonellasuplementado com ImmunoWall®0,5 kg/ton.
Gêneros marcantes aos 14 e 21 dias de idade:
Ruminococccus torques: Possui uma função mucolítica e é de especial interesse, pois está correlacionado com melhor produtividade em frangos (Torok et al., 2011). Sua presença foi menor no grupo desafiado com Salmonellado que no grupo controle. Embora não tenha sido avaliado, é interessante observar que o desafio pode ter afetado produtividade através da microbiota, e que o tratamento preveniu esta alteração. Subdoligranulum:Neste gênero foi detectada a expressão de butirato quinase, uma enzima envolvida no último passo de uma das quatro potenciais vias de produção de butirato (Polansky et al.,2016). O butirato é um ácido graxo de cadeia curta e desempenha um papel importante na fisiologia intestinal, servindo como fonte de energia para as células epiteliais do ceco e íleo, além de controlar bactérias patogênicas ao alterar o pH interno dos patógenos (Roto et al.,2015). A partir dessas características descritas, assume-se um papel positivo à bactériaSubdoligranulum, mas esse agente foi associado com pior conversão alimentar em aves (Singh et al., 2012). Assim sendo, a redução deste gênero pode ter sido um efeito positivo do tratamento, pois os resultados mostraram que ambos os tratamentos com ImmunoWall® reduziram a quantidade dessas bactérias.
Lactobacillus: Produz ácidos graxos de cadeia curta e produz bacteriocinas que impedem o crescimento de patógenos. Seus efeitos benéficos na saúde intestinal do hospedeiro já foram bem relatados, incluindo-se melhores respostas vacinais de anticorpos, melhor defesa contra Salmonellae maior produtividade (ganho de peso e eficiência alimentar) (Loman and Tappenden, 2016; Torok et al.,2011; Yan et al.,2017). Sabe-se que MOS e nucleotídeos de leveduras induzem o aumento de bactérias reconhecidamente benéficas, como Lactobacillus (Mesa et al., 2017). Confirmando a literatura, aos 14 dias ambos os tratamentos com ImmunoWall® aumentaram a proliferação de Lactobacillus.
Filos aos 21 dias de idade:
Proteobacteria: O tratamento com ImmunoWall® reduziu a quantidade deste filo em relação ao grupo controle. Tenericutes: Ambos os tratamentos com ImmunoWall® aumentaram a proliferação do filo Tenericutes aos 21 dias. Esse achado inédito pode ser de grande relevância visto que o gênero Tenericutes aparece em maior proporção no intestino de aves com melhor taxa de conversão alimentar (Singh et al.,2012).
A alteração benéfica encontradas na microbiota como o aumento de Lactobacillus e Tenericutes, pode responderpelas melhoras encontradas em outros sistemas.
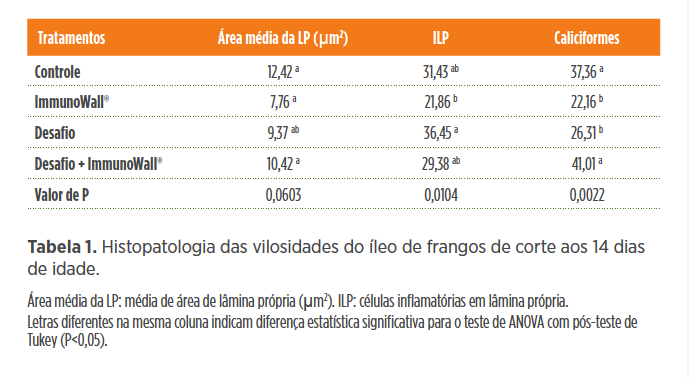
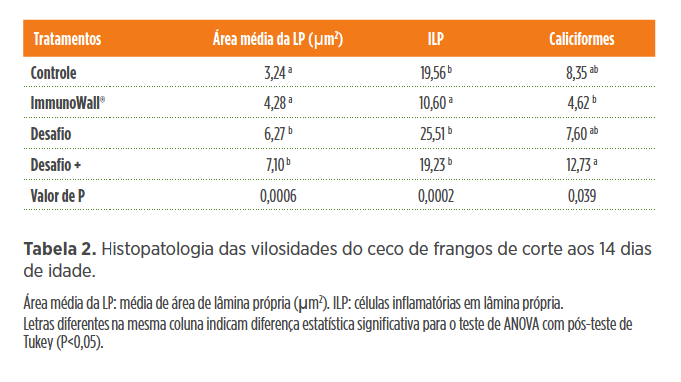
Essas bactérias podem potencializar as respostas imunes e melhorar a integridade intestinal. No mesmo estudo de Beirão et al.(2020, no prelo) foram avaliadas a área média da lâmina própria, células inflamatórias em lâmina própria e células caliciformes do íleo e ceco que permitem avaliar o tipo de resposta celular predominante frente ao agente patogênico (Tabelas 1 e 2). O ImmunoWall® melhorou a condição do intestino desafiado com SE, como podemos observar pela área da lâmina própria, onde se encontram grande parte das células imunes. A quantidade de células inflamatórias (ILP) foi reduzida com a suplementação de ImmunoWall® (Desafio vs. Desafio +ImmunoWall®;Controle vs. ImmunoWall®). As células caliciformes constituem uma das primeiras barreiras de defesa inespecífica do intestino e de acordo com os resultados verificamos que ImmunoWall® proporcionou maior produção nos animais desafiados. A suplementação de ImmunoWall® reduziu a quantidade de células inflamatórias (ILP) (Desafio vs. Desafio + ImmunoWall®; Controle vs.ImmunoWall®). Verificamos ainda que ImmunoWall® proporcionou maior produção de células caliciformes nos animais desafiados.
Os efeitos das leveduras sobre a microbiota não induzem sempre as mesmas alterações descritas, indicando que há variações importantes na condução dos testes, ambientes e produtos. Desta forma, esse efeito benéfico pode ser uma característica importante do ImmunoWall®.
Referências
Beirão B. C. B. et al. Yeast cell wall immunomodulatory and intestinal integrity effects on broilers challenged with Salmonella Enteritidis. Journal of Applied Poultry Research, 2020 (no prelo). Duda-Chodak, A., Tarko, T., Satora, P., Sroka, P., 2015. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: a review. European Journal of Nutrition. 54, 325–341. doi:10.1007/s00394-015-0852-y Flemming, J. S. Utilização de leveduras, probióticos e mananoligossacarídeos na alimentação de frangos de corte. 2005. 109 f. Tese (Doutorado em Tecnologia dos Alimentos) Universidade federal do Paraná, Curitiba, 2005. Gomes, M. O. S. Efeito da adição da parede celular de levedura sobre a digestibilidade, microbiota, ácidos graxos de cadeia curta e aminas fecais e parâmetros hematológicos e imunológicos de cães. 2009. 79f. Dissertação (Mestrado em Clinica Médica Veterinária). Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal, 2009. Huyghebaert, G., Ducatelle, R., Immerseel, F. Van, 2011. An update on alternatives to antimicrobial growth promoters for broilers. Veterinary Journal. 187, 182–188. doi:10.1016/j.tvjl.2010.03.003 Loman, B.R., Tappenden, K.A., 2016. Prebiotic Short-Chain Fructooligosaccharides (scFOS) Increases Abundance of the Butyrate Producing Microbial Community Differentially When Administered With or Without Probiotic Lactobacillus rhamnosus GG (LGG) in Piglets With Short-Bowel Syndrome (SBS). FASEB Journal. 30, 683–685. Mesa, D., Lammel, D.R., Balsanelli, E., Sena, C., Noseda, M.D., Caron, L.F., Cruz, L.M., Pedrosa, F.O., Souza, E.M., 2017. Cecal Microbiota in Broilers Fed with Prebiotics. Frontiers in Genetics. 8, 153. Petravić-Tominac, V. et al. Biological effects of yeast β-glucans. Agriculturae Conspectus Scientificus, n. 75, v. 4, 2010. Polansky, O., Sekelova, Z., Faldynova, M., Sebkova, A., Sisak, F., Rychlik, I., 2016. Important metabolic pathways and biological processes expressed by chicken cecal microbiota. Applied and Environmental Microbiology. 82, 1569–1576. Roto, S.M., Rubinelli, P.M., Ricke, S.C., 2015. An introduction to the avian gut microbiota and the effects of yeast-based prebiotic-type compounds as potential feed additives. Frontiers in Veerinary Science. 2, 28. Singh, K.M., Shah, T., Deshpande, S., Jakhesara, S.J., Koringa, P.G., Rank, D.N., Joshi, C.G., 2012. High through put 16S rRNA gene-based pyrosequencing analysis of the fecal microbiota of high FCR and low FCR broiler growers. Molecular Biology Reports. 39, 10595–10602. Torok, V.A., Hughes, R.J., Mikkelsen, L.L., Perez-Maldonado, R., Balding, K., MacAlpine, R., Percy, N.J., Ophel-Keller, K., 2011. Identification and Characterization of Potential Performance-Related Gut Microbiotas in Broiler Chickens across Various Feeding Trials. Applied and Environmental Microbiology. 77, 5868–5878. doi:10.1128/AEM.00165-11 Yan, W., Sun, C., Yuan, J., Yang, N., 2017. Gut metagenomic analysis reveals prominent roles of Lactobacillus and cecal microbiota in chicken feed efficiency. Scientific Reports. 7, 45308.
Por Liliana Borges e Melina Bonato (P&D, ICC Brazil)


 Uma microbiota saudável apresenta alta diversidade de gêneros microbianos em perfeito equilíbrio, permitindo um aumento da capacidade metabólica do intestino.
Uma microbiota saudável apresenta alta diversidade de gêneros microbianos em perfeito equilíbrio, permitindo um aumento da capacidade metabólica do intestino. A microbiota benéfica auxilia a digestão e absorção dos nutrientes, produz vitaminas que são utilizadas pelo hospedeiro e diminui, por competição exclusiva e/ou liberação de substâncias, a proliferação de agentes patogênicos. Esta microbiota nativa produz ácidos graxos de cadeia curta e ácido lático.
A microbiota benéfica auxilia a digestão e absorção dos nutrientes, produz vitaminas que são utilizadas pelo hospedeiro e diminui, por competição exclusiva e/ou liberação de substâncias, a proliferação de agentes patogênicos. Esta microbiota nativa produz ácidos graxos de cadeia curta e ácido lático. Com isso pode ocorrer uma melhoria nas estruturas e integridade das células, aumentando a capacidade de absorção dos nutrientes e consequente melhora no desempenho. Já as bactérias patogênicas causam inflamação na mucosa intestinal, geram metabólitos tóxicos, e propiciam o aparecimento de enfermidades,
Com isso pode ocorrer uma melhoria nas estruturas e integridade das células, aumentando a capacidade de absorção dos nutrientes e consequente melhora no desempenho. Já as bactérias patogênicas causam inflamação na mucosa intestinal, geram metabólitos tóxicos, e propiciam o aparecimento de enfermidades,  como é o caso da Salmonella, E. coli, Clostridium, Staphylococcus, Pseudomonas.
como é o caso da Salmonella, E. coli, Clostridium, Staphylococcus, Pseudomonas.


 A parede celular de levedura Saccharomyces cerevisiae (ImmunoWall®®, ICC Brazil) oriunda do processo de fermentação da cana-de-açúcar para produção de etanol, contém em torno de 35% de β-glucanas (1,3 e 1,6), e 20% de mananoligossacarídeos (MOS).
A parede celular de levedura Saccharomyces cerevisiae (ImmunoWall®®, ICC Brazil) oriunda do processo de fermentação da cana-de-açúcar para produção de etanol, contém em torno de 35% de β-glucanas (1,3 e 1,6), e 20% de mananoligossacarídeos (MOS).
 Um recente estudo de Beirão et al.(2018) onde frangos de corte foram alimentados com dietas suplementadas com ImmunoWall®® (0,5 kg/ton) e infectados aos dois dias de idade com SalmonellaEnteritidis [SE] (via oral na dosagem de 108UFC/ave), mostrou alterações na microbiota e histopatologia do íleo e ceco.
Um recente estudo de Beirão et al.(2018) onde frangos de corte foram alimentados com dietas suplementadas com ImmunoWall®® (0,5 kg/ton) e infectados aos dois dias de idade com SalmonellaEnteritidis [SE] (via oral na dosagem de 108UFC/ave), mostrou alterações na microbiota e histopatologia do íleo e ceco.
 positivo à bactériaSubdoligranulum, mas esse agente foi associado com pior conversão alimentar em aves (Singh et al., 2012).
positivo à bactériaSubdoligranulum, mas esse agente foi associado com pior conversão alimentar em aves (Singh et al., 2012). dessas bactérias.
dessas bactérias.