Intestinal inflammation and oxidative stress are becoming increasingly relevant to nutritionists and veterinarians working with monogastric animals. Any intestinal inflammation, even the sterile type produced by nutritional factors, results in an increase of oxidative stress. This renders further damage of intestinal mucosa, expresed as reduced feed intake, impaired body weight gain, and diarrhoea. This article reviews the mechanisms involved in the development of enteritis, as well as its relationship with oxidatve stress.
Introduction
Currently, the term “intestinal health” is no doubt one of the most used expressions in the nutrition of monogastric animals, in both scientific and commercial/marketing areas. We could assume that it is simply a buzzword, however the intestine and its health status play a major role in the appropriate physiological development of these animals and, ultimately, in their growth and productive performance.
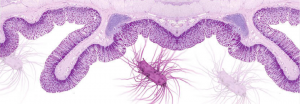
Intestinal mucosa
The intestinal epithelium is the largest surface area of the body in contact with the external environment. It acts as an effective barrier, reducing the exposure to certain harmful environmental elements and pathogens, allowing for nutrient absorption and secretion of waste substances.
Also, the intestinal environment provides a platform for the growth of a diverse microbiota. This community of microorganisms is not only a second barrier against pathogen colonization, but also it regulates the immune development, and provides metabolites for the nutrition of the host.
Finally, the intestine has neuroendocrine functions,participating in the regulation of many physiological functions of the animal.
Any imbalance between these components and their functions will generate an intestinal inflammation, as an ubiquitous defence mechanism of monogastric animals. The intestinal inflammation is the pathogenic basis of all enteric disorders (clinical or subclinical).
Figure 1. Oxidative stress as a result of an imbalance between ROS production and endogenous antioxidant activity.
Oxidative stress
In recent years, another term has arisen in the vocabulary of the animal health and nutrition sector. This term, which is becoming more popular through many publications and presentations, is known as “oxidative stress”.
As we know, oxygen is necessary for animal life since it acts as the final acceptor of four electrons in the mitochondrial respiration. However, when the oxygen is partially reduced, reactive oxygen species (ROS) are generated. These are molecules containing an oxygen atom with at least one unpaired electron.
Figure 1. Oxidative stress as a result of an imbalance between ROS production and endogenous antioxidant activity.
ROS are usually very reactive and able to capture electrons from other nearby molecules, including DNA, lipids, and proteins. The consequent changes to the structure of these macromolecules cause serious metabolic disorders and tissue damage.
Under normal conditions, there is a balance in the generation of ROS and the endogenous antioxidant defence system.
However, small changes in metabolism or immune response can upset this delicate balance, generating an oxidative stress. Therefore, the oxidative stress condition is characterized by excessive ROS production and/or insufficient protection by the endogenous antioxidant defense system (Fig. 1).
Not all intestinal inflammations are the same
In pig and poultry farming, animals are continuously exposed to countless potentially pathogenic agents, through the air, water, feed, and contact with other animals. This results in the activation of the intestinal innate immune system.
Intestinal inflammation is the response of the system to those agents and to the modifications in tissue homeostasis caused by them.
The starting points of the intestinal inflammation are numerous immune receptors called PRRs (pattern recognition receptors) found in the intestinal epithelium and in the lamina propria. They detect molecules associated with microbial pathogens, as well as other danger signs – non-microbial – or cell debris derived from epithelial damage. The recognition activates a highly regulated network of immunological and physiological processes to maintain homeostasis. The result is the production of a variety of proinflammatory cells and molecules, essential to control most of the enteric challenges faced by the animals.
Types of enteric inflammation in monogastric animals
Figure 2. Types of inflammation
The intestinal inflammation is an adaptive response that adjusts to the type of trigger that has caused it. Three types of intestinal inflammations have been identified, namely physiological, pathological, and sterile inflammations.
Physiological inflammation
Even under balanced conditions and good good intestinal health, the gut mucosa is continuously exposed to several exogenous antigenic stimulation from the feed. There is also an endogenous stimulation by certain bacterial structural components (lipopolysaccharides and peptidoglycans) from the commensal microbiota, or metabolites derived from them. The interaction between PRR receptors in the intestine and such stimuli maintains a constant mild activation of the innate immune system without inducing disease. That is known as “physiological inflammation”.
This type of low-grade inflammation has been described both in poultry (Kogut et al., 2018) and pigs (Splichalova et al., 2018). It is a controlled inflammatory response based on the balance between defense capacity and tolerance of the innate immune system with regards to the intestinal microbiota.
Pathological inflammation
In cartain situations, the defense- tolerance balance of the innate immune system is disrupted, transforming the physiological inflammation into a more severe one, known as “pathological inflammation” or “classical inflammation”. That disruption can result from the occurrence of an intestinal acute infectious process, or tissue damage. The consequent increase in the concentration of specific molecules of pathogenic microorganisms, such as virulent factors, or the presence of debris from damage tissue, trigger the mentioned imbalance.
Sterile inflammation
However, it is important to note that, in the absence of infection, a chronic inflammation can occur in response to certain chemical/ physical substances and metabolic stimulation, known as “sterile inflammation”(Rubartelli et al., 2013; Kogut et al., 2018; Splichalova et al., 2018).
Certain ingredients used in animal feeds can contain components that trigger the sterile inflammation.
Soybean meal contains many antinutritional factors likely to cause inflammation in the intestinal epithelium of monogastric animals. Such factors include the allergen β-conglycinin or 75 globulin (Chen et al., 2011; Penget al., 2018).
The β-conglycinin, found in soybean meal, causes an increase in proinflammatory cytokines in the intestine (Peng et al., 2018). They also cause direct damage of the gut epithelium, depressed cell growth, disrupted cytoskeleton, and cellular apoptosis (Chen et al., 2011).
Prolamins, which are part of the gluten found in cereals such as corn and wheat, can also cause chronic sterile inflammation. They alter the expression of the proteins forming the tight junctions between adjacent enterocytes, increasing intestinal permeability. This can contribute to the induction of sterile inflammation (Punder and Pruimboom, 2015).
Other triggers of sterile inflammation of dietary origin are lipids with a certain level of rancidity. They are important sources of ROS, which induce the development of mucosal inflammation. Therefore, the quality of fats and oils added to animal feeds, as well as the quality of raw materials of animal origin rich in lipids (e.g. as fish meal), may play a significant role in the development of sterile inflammation (Kogut et al., 2018; Mavromichalis, 2019).
In short, the sterile inflammation of dietary origin can be just as harmful as the pathological inflammation caused by microbial infections, since the final results are the same:
loss of appetite
reduction of productive performance
diarrhea, or even death, due to secondary complications caused by opportunistic microorganisms.
The oxidative stress damages cells
Several factors can trigger oxidative stress in the animals, such as:
high metabolic activity, as typically seen in the intensive production systems
stressful times, such as the weaning of piglets heat (particularly in broilers)
heat (particularly in broilers)
diet
These are situations where oxygen requirements are increased and, therefore, either more oxidation occurs or elements (diet) that introduce oxidizing agents in the organism are produced.
These are situations where there is an increment in the oxygen needs, consequently producing more oxidation, or where exogenous oxidising agents enter the animal though the diet (for example).
During the oxidative stress, macromolecules (particularly DNA, proteins, and lipids) are the natural targets of ROS. As described by Poulsen (2005), ROS can interact with the DNA, producing numerous modifications that can result in mutations, double-strand breaks in DNA and, finally, cell death.
Protein oxidation results in abnormal enzymes that do not work properly and are unable to perform their cellular tasks. All the metabolic pathways in which they participate are altered.
Lipids are important constituents of the cell membrane lipid bilayer. Unsaturated fatty acids are particularly susceptible to oxidation. Oxidative chain reactions may occur, causing damage that can compromise cell integrity.
In this process, the extraction of a hydrogen atom by ROS produces a conjugated diene, making the lipid more susceptible to additional oxidation. Its subsequent reaction with the molecular oxygen results in the formation of a lipid peroxyl radical that is capable of oxidizing other lipids, spreading the oxidative damage (Lykkesfeldt and Svendsen, 2007).
In view of the damage caused by oxidative stress, the mechanisms of cellular defense can be divided into at least three levels, according to the following functions:
neutralize the oxidants
repair the damage
encapsulate when no repair is possible
Inflammation and oxidative stress: a vicious cycle
The inflammatory processes are always associated with oxidative stress.
During mucosal inflammation, the intestinal epithelial cells, as well as neutrophils (heterophils in birds) and macrophages, produce superoxide (a ROS) and nitric oxide (a reactive nitrogen species, or RNS). This occurs when the oxidative enzymes NADPH oxidase (NOX) and nitric oxide synthase (iNOS), respectively, are induced by proinflammatory cytokines secreted as part of the inflammatory process (Tian et al., 2017).
Any type of intestinal inflammation will generate ROS and NO and, therefore, will lead to intestinal oxidative stress.
On the other hand, the NO produced during the inflammatory process also has antimicrobial properties. When released in the intestinal lumen, the nitric oxide (NO) is rapidly converted to nitrate. The growth of E. coli can benefit from this nitrate- rich environment found in the inflamed intestine, since this pathogen has nitrate-reducing genes. Also, the increased blood flow in the inflamed intestine enhances the oxygen concentration, which can result in an explosion of growth of facultative anaerobes. These bacteria are common pathogenic microorganisms, such as species of the genus Streptococcus and Enterobacteriaceae (E. coli). Therefore, the occurrence of inflammation and the subsequent oxidative stress can result in the increase of certain populations of pathogenic bacteria and the reduction of non-pathogenic bacterial populations.
The condition of microbial imbalance in the intestine is called «dysbiosis» and leads to a loss of intestinal microbial diversity.
This dysbiosis derives from any type of inflammation, including the sterile inflammation, and may result in pathological inflammation in pigs and poultry, as indicated by Lauridsen (2018).
Also, the oxidative stress associated with inflammation can modify the phenotype of the intestinal epithelial cells, which causes changes in the expression of certain genes. For example, under conditions of intestinal oxidative stress, there is an up-regulation of the expression of proinflammatory molecules such as IL-8 and IL-1, transforming growth factor beta (TGF beta). There is also a down-regulation of occludin and mucin 2, which are responsible for the integrity of intestinal epithelium (Ducatelle et al. 2018).
Therefore, the oxidative stress generated by sterile and pathological intestinal inflammation increases their severity, entering a considerable vicious cyle that will lead to a chronic inflammatory condition (Figure 4). In the past, the proven anti- inflammatory action of certain antibiotic growth promoters impeded the establishment of such vicious circle.
In conclusion
Any enteric inflammation, including the sterile inflammation caused by dietary factors, results in intestinal oxidative stress. Also, this oxidative stress induces the gene expression of different proinflammatory molecules, leading to an even higher degree of inflammation.
Consequently, controlling the introduction of pro-inflammatory factors or oxidant elements through the feed may be a decisive tactic to minimize the incidence of enteric disorders in monogastric animals.