The intestine is considered as the the largest immune organ in the body.
The intestinal structure is crucial for the health and performance of animals. In addition to the digestive functions of the gastrointestinal tract (GIT), the immune cells and lymphoid structures present in the GIT constitute the largest immune organ in the body.
The foundations of the mucosal-associated immune system in the intestine are divided into five spheres, which are:
![]() Epithelial barrier, a physical barrier composed of epithelial cell layers (enterocytes and colonocytes), mucus-producing goblet cells (goblet cells), bactericidal substance-producing Paneth cells (defensins), carrier cells (M cells), and intraepithelial T lymphocytes (Figure 1).
Epithelial barrier, a physical barrier composed of epithelial cell layers (enterocytes and colonocytes), mucus-producing goblet cells (goblet cells), bactericidal substance-producing Paneth cells (defensins), carrier cells (M cells), and intraepithelial T lymphocytes (Figure 1).
![]() Isolated lymphoid follicles, site of initiation of adaptive immune responses, organized structure of immune cell populations.
Isolated lymphoid follicles, site of initiation of adaptive immune responses, organized structure of immune cell populations.
![]() Peyer’s patches, the site of initiation of adaptive immune responses, an organized structure of multiple lymphoid follicles with germinal centers.
Peyer’s patches, the site of initiation of adaptive immune responses, an organized structure of multiple lymphoid follicles with germinal centers.
![]() Diffuse lymphoid tissue, macrophages, dendritic cells, mast cells, irritating T lymphocytes, regulatory T cells, IgA-secreting plasma cells scattered within the lamina propia of the mucosa.
Diffuse lymphoid tissue, macrophages, dendritic cells, mast cells, irritating T lymphocytes, regulatory T cells, IgA-secreting plasma cells scattered within the lamina propia of the mucosa.
![]() Mesenteric lymph nodes, the site of initiation of adaptive immune responses against intestinal antigens brought by the lymphatic system (Gonçalves et al., 2016).
Mesenteric lymph nodes, the site of initiation of adaptive immune responses against intestinal antigens brought by the lymphatic system (Gonçalves et al., 2016).
 Figure 1. Structure of defense of the intestinal epithelial barrier (adapted from D. Artis. 2008; 8:411 Nature Reviews Immunology).
Figure 1. Structure of defense of the intestinal epithelial barrier (adapted from D. Artis. 2008; 8:411 Nature Reviews Immunology).
Intestinal surface
The surface of the mucosa (Figure 2) is covered with mucus formed by mucins, which are secreted by goblet cells and create a barrier that prevents larger particles, including most bacteria, from coming into direct contact with the layer of epithelial cells (Turner, 2009).

Mucins contain different oligosaccharides and include secreted and cell surface glycoproteins. The secreted mucins, including MUC2, MUC5, and MUC6, form a hydrated gel layer that is 300 to 700 μm thick and consists of two layers: an outer less dense layer that is typically colonized by bacteria, and an inner dense layer that is attached to the epithelium and is free of bacteria.
 Figure 2. Representation of the intestinal mucosal epithelium.
Figure 2. Representation of the intestinal mucosal epithelium.
Paneth cells (Figure 3), located in the crypts of the small intestine, secrete α-defensins. In the colon, β-defensin peptides are produced by absorptive epithelial cells in intestinal crypts, some constitutively and others in response to the proinflammatory cytokine IL-1 (Abbas et al., 2015).
![]() The expression of regenerating islet-derived protein III gamma (REGIIIγ) by intestinal epithelial cells requires signals from Toll-like receptors (TLRs) in response to invading organisms, and its production increases after colonization and infection by pathogens (Abbas et al., 2015).
The expression of regenerating islet-derived protein III gamma (REGIIIγ) by intestinal epithelial cells requires signals from Toll-like receptors (TLRs) in response to invading organisms, and its production increases after colonization and infection by pathogens (Abbas et al., 2015).
 Figure 3. Paneth cell.
Figure 3. Paneth cell.

Immune response
![]()
The innate immune response begins with the mediation through the recognition of pathogen-associated molecular patterns (PAMPs), as well as cellular pattern recognition receptors (PRRs). Toll-like receptors (TLRs) function as PRRs in mammals and play a crucial role in recognizing microbial components and initiating the innate immune response.
TLRs are divided into two subgroups, depending on their cellular location and specificity regarding their respective PAMPs.
![]() One group consists of TLR-1, TLR-2, TLR-4, TLR-5, TLR-6, and TLR-10, which are expressed on cell surfaces and recognize microbial membrane components such as lipids, lipoproteins, and proteins.
One group consists of TLR-1, TLR-2, TLR-4, TLR-5, TLR-6, and TLR-10, which are expressed on cell surfaces and recognize microbial membrane components such as lipids, lipoproteins, and proteins.
 This reaction can trigger an immune response, which can lead to an exaggerated and pathological immune response, culminating in a process of intestinal inflammation (Gonçalves et al., 2016).
This reaction can trigger an immune response, which can lead to an exaggerated and pathological immune response, culminating in a process of intestinal inflammation (Gonçalves et al., 2016).
 Figure 4. Immediate immune system response to pathogen invasion.
Figure 4. Immediate immune system response to pathogen invasion.
Under these conditions, for effective protection, the animal organism must possess defense systems that detect and eliminate invading microorganisms efficiently, preferably without causing tissue damage and discomfort, a function attributed to the adaptive immune system (Levinson, 2016).
![]()
Adaptive immune responses in the gastrointestinal tract (GIT) are initiated in discretely organized sets of lymphocytes and antigen-presenting cells closely associated with the epithelial mucosal lining of the intestine, as well as in the mesenteric lymph nodes (Abbas et al., 2015).
![]() Organized and non-encapsulated lymphoid tissues consisting of immune cells are associated with mucosal surfaces (Figure 5) of the respiratory, gastrointestinal, and urogenital tracts, collectively referred to as MALT (Mucosa-associated lymphoid tissue).
Organized and non-encapsulated lymphoid tissues consisting of immune cells are associated with mucosal surfaces (Figure 5) of the respiratory, gastrointestinal, and urogenital tracts, collectively referred to as MALT (Mucosa-associated lymphoid tissue).
![]() Specifically in the mucosa of the GIT, there is the GALT system (gut-associated lymphoid tissue), which consists of dense lymphoid tissue (Gonçalves et al., 2016).
Specifically in the mucosa of the GIT, there is the GALT system (gut-associated lymphoid tissue), which consists of dense lymphoid tissue (Gonçalves et al., 2016).

Figure 5. Organization of lymphoid tissues on the surface of the intestinal tract.
In GALT, lymphoid tissue is distributed along structures such as:
![]() Mesenteric lymph nodes (LN),
Mesenteric lymph nodes (LN),
![]() Lamina propria (LP), such as Peyer’s patches (PP) in the ileum, or as more diffuse lymphoid aggregates (Vega-Lopez et al., 2012).
Lamina propria (LP), such as Peyer’s patches (PP) in the ileum, or as more diffuse lymphoid aggregates (Vega-Lopez et al., 2012).
In addition to these structures, antigen-presenting cells (APCs), dendritic cells (DCs), macrophages, T cells, and B cell areas with germinal centers in the LP, and natural killer (NK) cells form the GALT structure (Cunha, 2013).
The most prominent GALT structures are Peyer’s patches, which are primarily found in the distal ileum, and in small aggregates of lymphoid follicles or isolated follicles in the appendix and colon.
![]()
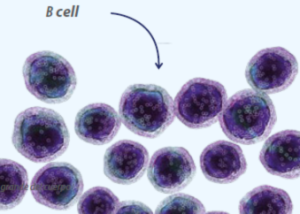 Peyer’s patches have the structure of lymphoid follicles, with germinal centers containing B lymphocytes, follicular helper T cells, follicular dendritic cells, and macrophages. The germinal centers within the follicles are surrounded by immature follicular B cells that express IgM and IgD.
Peyer’s patches have the structure of lymphoid follicles, with germinal centers containing B lymphocytes, follicular helper T cells, follicular dendritic cells, and macrophages. The germinal centers within the follicles are surrounded by immature follicular B cells that express IgM and IgD.
A region called the dome located between the follicles and the lining epithelium contains B and T lymphocytes, dendritic cells, and macrophages (Abbas et al., 2015).
The main pathway for antigen distribution within GALT occurs through specialized cells called microfold cells (M cells), which are found in regions of the intestinal epithelium called dome epithelium or follicle-associated epithelium (FAE) (Abbas et al., 2015).

These cells have unique morphological characteristics, including reduced glycocalyx presence, irregular brush border, and reduced microvilli. It is worth noting that M cells are highly specialized in the phagocytosis and transcytosis of macromolecules from the intestinal lumen, particulate antigens, and pathogenic or commensal microorganisms across the epithelium (Mabbott et al., 2013).
While M cells play a crucial role in the immune response against luminal microorganisms, some pathogenic microorganisms have evolved to deceive the M cell mechanism of action, using them as a pathway for invasion through the mucosal barrier.
The best-described example is Salmonella typhimurium, similar to the human pathogen S. typhi, which causes typhoid fever. M cells express lectins that allow specific binding and internalization of these bacteria.
![]() Bacteria are cytotoxic to M cells, leading to increased intestinal permeability (Figure 6) and subsequently the proliferation of microorganisms (Abbas et al., 2015).
Bacteria are cytotoxic to M cells, leading to increased intestinal permeability (Figure 6) and subsequently the proliferation of microorganisms (Abbas et al., 2015).

Figure 6. Some pathogens use M cells as a pathway for invasion through the mucosa. Subsequently, other defense cells come into action.
Sequence of the inflammatory response
Dendritic cells and macrophages are sentinel cells and antigen processors; as a result, antigen processing can be initiated simultaneously with the elimination of the invader by innate defenses.
![]()
![]() After phagocytosis, the invading microorganisms will be processed intracellularly and fragmented into smaller peptides, which will then bind to specialized antigen-presenting receptors called major histocompatibility complex (MHC) molecules and be transported to the cell surface, so that the triggering of adaptive immunity occurs when these MHC-bound peptides are recognized by specific receptors on lymphocytes (Tizard, 2008).
After phagocytosis, the invading microorganisms will be processed intracellularly and fragmented into smaller peptides, which will then bind to specialized antigen-presenting receptors called major histocompatibility complex (MHC) molecules and be transported to the cell surface, so that the triggering of adaptive immunity occurs when these MHC-bound peptides are recognized by specific receptors on lymphocytes (Tizard, 2008).
Defense in circulation
 There are also effector T and B lymphocytes (helper CD4 T lymphocytes, cytotoxic CD8 T lymphocytes, and IgA-secreting plasma cells) previously activated/differentiated in GALT or draining lymph nodes (Gonçalves et al., 2016).
There are also effector T and B lymphocytes (helper CD4 T lymphocytes, cytotoxic CD8 T lymphocytes, and IgA-secreting plasma cells) previously activated/differentiated in GALT or draining lymph nodes (Gonçalves et al., 2016).
These lymphocytes are heterogeneous, mainly (80%) CD8 phenotype, with abundant cytoplasmic granules containing cytotoxic molecules, the ability to produce various cytokines (such as IFN-γ, IL-2, IL-4, or IL-17), and can be divided into cellular populations that express on the surface the antigen receptor (TCR/T cell receptor) composed of αβ or γδ chains (Gonçalves et al., 2016).

The dominant intraepithelial T lymphocytes correspond to CD8αβ+/TCRαβ+, which penetrate the intestinal epithelium, increasing the expression of specific integrins and chemokine receptors after activation in secondary lymphoid organs (Gonçalves et al., 2016).
Natural intraepithelial T lymphocytes TCRγδ+ have primary functions in the intestinal mucosa, including:
![]() Maintenance of epithelial barrier function (control of epithelial cell growth, secretion of TGF-β),
Maintenance of epithelial barrier function (control of epithelial cell growth, secretion of TGF-β),
![]() Tissue Homeostasis
Tissue Homeostasis
![]() Protection against intestinal pathogens (production of inflammatory cytokines, cytotoxicity) (Gonçalves et al., 2016).
Protection against intestinal pathogens (production of inflammatory cytokines, cytotoxicity) (Gonçalves et al., 2016).
T lymphocytes are essentially CD4+ or CD8+ (both TCRαβ+), exhibit a memory phenotype (CD45RD), and are associated with the intestinal mucosa through the expression of specific integrins (α4β7) and chemokine receptors (CCR9) that direct these lymphocytes after their activation and differentiation into effector cells (Gonçalves et al., 2016).
 For the adaptive immune response to enteric antigens to occur, it is necessary for intravascular naive T lymphocytes (T naïve) to migrate to GALT and draining mesenteric lymph nodes, where they will be activated through clonal expansion, polarization/differentiation into Th1 and/or Th17 effector cells upon initial interaction with the antigen-presenting cell.
For the adaptive immune response to enteric antigens to occur, it is necessary for intravascular naive T lymphocytes (T naïve) to migrate to GALT and draining mesenteric lymph nodes, where they will be activated through clonal expansion, polarization/differentiation into Th1 and/or Th17 effector cells upon initial interaction with the antigen-presenting cell.
Subsequently, effector T lymphocytes exit the lymphoid tissue through efferent lymphatic vessels, enter the systemic bloodstream, and return to the intestine, where they will assist in the elimination of a specific antigen (Gonçalves et al., 2016), triggering the immune response, which in turn is already amplified and has a more diverse population of antigen-presenting cells, such as:
![]() Macrophages,
Macrophages,
![]() B lymphocytes,
B lymphocytes,
![]() In addition to dendritic cells.
In addition to dendritic cells.
In this new contact with the antigen (promoted by antigen-presenting cells), effector T lymphocytes respond more rapidly and vigorously, secreting cytokines such as IFN-γ, IL-17, TNF-α, lymphotoxin-α, or IL-2, depending on the profile of the effector T cell (Th1 or Th17). Each cytokine has a specific function in coordinating the triggered immune response.
IFN-γ stimulates antigen-presenting cells to produce IL-12 and specifically activates the production of other inflammatory cytokines such as IL-1, IL-6, IL-8, IL-18, and TNF-α, as well as reactive oxygen and nitrogen species in macrophages (Gonçalves et al., 2016).
![]()
IL-2 stimulates the growth and proliferation of T lymphocytes and B cells, as well as inducing the production of other cytokines, such as IFN-γ and TNF-β, resulting in the activation of monocytes, neutrophils, and natural killer cells (Oliveira et al., 2011).
 B lymphocytes play a central role in the immune response of mucosal membranes through antibody production.
B lymphocytes play a central role in the immune response of mucosal membranes through antibody production.
These cells are activated in mesenteric lymph nodes, isolated lymphoid follicles of the intestinal mucosa, and also in Peyer’s patches, with the cytokine TGF-β, and differentiate into antibody-producing cells of the IgA isotype in its dimeric form, secretory IgA (sIgA), or polymeric IgA.
![]() IgA plays an important role in protecting the mucosal epithelium against invasive pathogens, modulating the composition of the intestinal microbiota, and maintaining homeostasis against commensal and dietary antigens (Gonçalves et al., 2016).
IgA plays an important role in protecting the mucosal epithelium against invasive pathogens, modulating the composition of the intestinal microbiota, and maintaining homeostasis against commensal and dietary antigens (Gonçalves et al., 2016).
![]() IgAs bind to commensal bacteria in the intestinal lumen and allow their transport to the mucosa through the binding of the sIgA/antigen complex to specific receptors expressed on intestinal M cells.
IgAs bind to commensal bacteria in the intestinal lumen and allow their transport to the mucosa through the binding of the sIgA/antigen complex to specific receptors expressed on intestinal M cells.
This process of sampling commensal microorganisms to immune cells in a regulatory (anti-inflammatory) microenvironment allows the maintenance of tolerance to the intestinal microbiota (Gonçalves et al., 2016).
In view of the above, it is evident that the intestine, in addition to being an organ responsible for the processes of digestion and nutrient absorption, plays a crucial role in the defense and immune response of the body. Thus, the maintenance of intestinal health will be reflected in the health and productive performance of animals.
You may also like to read: “Immunomodulatory and immunostimulating effects of beta-glucans.”