06 Sep 2024
06 Sep 2024
Boletín técnico para MiaBond 360 en ganadería lechera
 Las micotoxinas son metabolitos secundarios que generan los hongos y mohos que infestan las plantas y sus productos, tanto durante la fase vegetativa como en el almacenamiento (García, 2023).
Las micotoxinas son metabolitos secundarios que generan los hongos y mohos que infestan las plantas y sus productos, tanto durante la fase vegetativa como en el almacenamiento (García, 2023).
Los rumiantes parecen ser bastante tolerantes de los efectos adversos de las micotoxinas y tal vez ello se deba a la capacidad de la microflora del rumen para detoxificarse.
 Sin embargo, se cree que las vacas lecheras de alta producción son más sensibles a la micotoxicosis, ya que ingieren cantidades más elevadas de pienso.
Sin embargo, se cree que las vacas lecheras de alta producción son más sensibles a la micotoxicosis, ya que ingieren cantidades más elevadas de pienso.
Esta mayor ingesta provoca un paso más rápido por el tracto digestivo y, en consecuencia, un menor tiempo para la detoxificación de las micotoxinas por parte de los microorganismos del rumen (Nährer, 2015; Seglar, 2024).

Los aglutinadores de micotoxinas son aditivos del pienso utilizados en el control de las micotoxinas. Actúan adhiriéndose, neutralizando e impidiendo que las micotoxinas lleguen a entrar en el torrente sanguíneo (Yildiz, 2024).
 MiaBond 360 es más que un aglutinador de toxinas. Contiene bentonita, polifenoles, enzima fumonisina esterasa y bacterias de la familia Coriobacteriaceae.
MiaBond 360 es más que un aglutinador de toxinas. Contiene bentonita, polifenoles, enzima fumonisina esterasa y bacterias de la familia Coriobacteriaceae.
![]() Por lo tanto, MiaBond 360 proporciona una protección integral contra las micotoxinas y los problemas de salud asociados con las mismas.
Por lo tanto, MiaBond 360 proporciona una protección integral contra las micotoxinas y los problemas de salud asociados con las mismas.
La aflatoxina es una micotoxina con frecuencia presente en los piensos para animales (Jiang et al., 2019). La mayor parte de la normativa se refiere al control de la aflatoxina, ya que se considera la más tóxica y carcinogénica de las micotoxinas naturales (Whitaker y Slate, 2015).
![]() Las bentonitas han mostrado los resultados más prometedores en lo que respecta a la adsorción de la aflatoxina B1 (Rodrigues, Nährer y Schwab, 2015).
Las bentonitas han mostrado los resultados más prometedores en lo que respecta a la adsorción de la aflatoxina B1 (Rodrigues, Nährer y Schwab, 2015).
No todas las bentonitas son iguales. Por ejemplo, la capacidad adsortiva de la aflatoxina B1 es variable.
MiaBond 360 incorpora bentonita aprobada por la Unión Europea. Por lo tanto, cumple con todos los criterios requeridos para una elevada capacidad aglutinadora.
![]() La capacidad de adsorción no depende del nivel de pH del entorno. El nivel de pH de un rumen saludable oscila entre 6,2 y 6,4. Sin embargo, los niveles del pH cambian en las partes más bajas del sistema digestivo y pueden descender hasta 2 en el abomaso.
La capacidad de adsorción no depende del nivel de pH del entorno. El nivel de pH de un rumen saludable oscila entre 6,2 y 6,4. Sin embargo, los niveles del pH cambian en las partes más bajas del sistema digestivo y pueden descender hasta 2 en el abomaso.
Eso implica que si el aglutinador de toxinas no incluye bentonita de alta calidad, la mayor parte de las moléculas adsorbidas de aflatoxina serán liberadas en el abomaso. Posteriormente serán absorbidas a través del intestino y entrarán en el torrente sanguíneo, acarreando posibles problemas de salud y rendimiento de la vaca lechera.
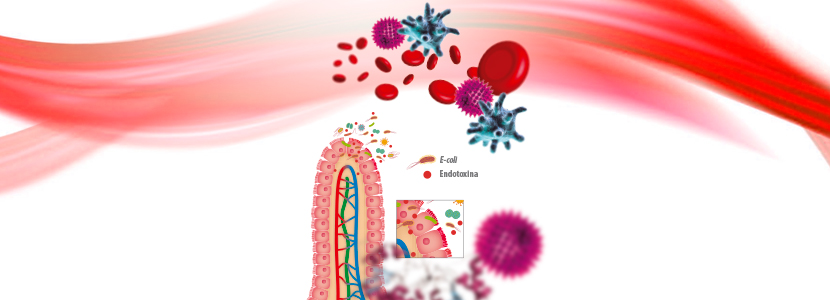
Las dietas ricas en cereales en vacas lecheras están asociadas a una rápida disminución del pH del rumen, produciendo cambios significativos en su población microbiana.
Hay cada vez más pruebas de que estas dietas también están vinculadas a la liberación de grandes cantidades de endotoxinas en el líquido ruminal.
Las endotoxinas, también denominadas «lipopolisacáridos» (LPS), son los componentes principales de las membranas de la pared celular de las bacterias gram-negativas (BGN). No son perjudiciales cuando están dentro de la membrana celular, pero cuando se liberan, producen un efecto altamente inmunoestimulador y son responsables en gran medida de los signos de infección bacteriana.
![]() A medida que se incrementa la proporción de carbohidratos no fibrosos en la dieta, se observa un aumento consecuente de las BGN, mientras que la aparición de acidosis ruminal subaguda (ARSA) causa una reducción importante de la población de BGN.
A medida que se incrementa la proporción de carbohidratos no fibrosos en la dieta, se observa un aumento consecuente de las BGN, mientras que la aparición de acidosis ruminal subaguda (ARSA) causa una reducción importante de la población de BGN.
La disminución de las BGN podría estar asociada a un incremento de los LPS ruminales libres, probablemente debido a la muerte y lisis de las bacterias. Las condiciones de ARSA pueden resultar no solo en el lisado de las BGN, sino también afectar negativamente a la integridad de las paredes del rumen.
 En este caso, la ubre y el útero pueden asimismo convertirse en fuentes importantes de endotoxinas.
En este caso, la ubre y el útero pueden asimismo convertirse en fuentes importantes de endotoxinas.
 Hay cada vez más pruebas de que las endotoxinas se trasladan del rumen, útero y ubres a la circulación sistémica, induciendo afecciones inflamatorias crónicas de grado bajo que están asociadas a múltiples enfermedades, entre ellas hígado graso, mastitis, placenta retenida, metritis, laminitis, abomaso desplazado, fiebre láctea y síndrome de la vaca decaída (Andersen, 2003; Ametaj, Zebeli y Iqbal, 2010; Chodorowska, 2016; Eckel y Ametaj, 2016; Eckel y Ametaj, 2020).
Hay cada vez más pruebas de que las endotoxinas se trasladan del rumen, útero y ubres a la circulación sistémica, induciendo afecciones inflamatorias crónicas de grado bajo que están asociadas a múltiples enfermedades, entre ellas hígado graso, mastitis, placenta retenida, metritis, laminitis, abomaso desplazado, fiebre láctea y síndrome de la vaca decaída (Andersen, 2003; Ametaj, Zebeli y Iqbal, 2010; Chodorowska, 2016; Eckel y Ametaj, 2016; Eckel y Ametaj, 2020).
![]() Se ha demostrado sólidamente la asociación de la acidosis ruminal con la laminitis.
Se ha demostrado sólidamente la asociación de la acidosis ruminal con la laminitis.
![]() El mecanismo por el que se desarrolla la laminitis está ligado a la acidosis, mediante la depresión del pH sistémico, el cual activa un mecanismo vasoactivo que incrementa la presión sanguínea y que resulta exacerbado por factores como las histaminas y endotoxinas (Nocek, 1997).
El mecanismo por el que se desarrolla la laminitis está ligado a la acidosis, mediante la depresión del pH sistémico, el cual activa un mecanismo vasoactivo que incrementa la presión sanguínea y que resulta exacerbado por factores como las histaminas y endotoxinas (Nocek, 1997).
La cojera está asociada a un número más alto de servicios por concepción y, en consecuencia, a tasas de concepción a primer servicio más bajas (Andersen, 2003; Meléndez et al., 2003; Ametaj, Zebeli e Iqbal, 2010).
La mortalidad embrionaria temprana y su elevada frecuencia en vacas lactantes es una de las causas de la reducción del rendimiento y reproducción animal, y de la efectividad de la industria moderna de la ganadería lechera en su conjunto (Nezhdanov et al., 2017).
![]() Un evento de este tipo en teoría no tiene ningún efecto sobre la duración del ciclo estrogénico, y explica hasta el 70 %-80 % de las pérdidas de embarazo totales (BonDurant, 2004). Cullor (1993) observó que los LPS inducían luteolisis, y una reducción de la tasa de concepción y de la supervivencia embrionaria temprana a través de la liberación de mediadores inflamatorios.
Un evento de este tipo en teoría no tiene ningún efecto sobre la duración del ciclo estrogénico, y explica hasta el 70 %-80 % de las pérdidas de embarazo totales (BonDurant, 2004). Cullor (1993) observó que los LPS inducían luteolisis, y una reducción de la tasa de concepción y de la supervivencia embrionaria temprana a través de la liberación de mediadores inflamatorios.
Como se ha indicado anteriormente, las endotoxinas tienen enormes efectos negativos sobre el ganado vacuno lechero.
No obstante, tenemos la oportunidad de contrarrestar las endotoxinas mediante el uso de un aglutinador de toxinas complejo como MiaBond 360.
 La bentonita incluida en MiaBond 360 tiene una gran capacidad de aglutinación de las endotoxinas, superior al 99 %.
La bentonita incluida en MiaBond 360 tiene una gran capacidad de aglutinación de las endotoxinas, superior al 99 %.
Las micotoxinas pueden incrementar la producción celular de especies de oxígeno reactivo (EOR). La exposición a condiciones que potencian el estrés oxidativo podría comprometer la capacidad del embrión bovino para desarrollarse hasta el estadio de blastocito previamente a la implantación (Jiang et al., 2019).
 Los efectos adversos del estrés oxidativo sobre el embrión no solo están asociados a la embriotoxicidad directa de los agresivos radicales libres, sino también a su efecto negativo sobre las estructuras de síntesis hormonal de las gónadas y del sistema inmunitario (Nezhdanov et al., 2017).
Los efectos adversos del estrés oxidativo sobre el embrión no solo están asociados a la embriotoxicidad directa de los agresivos radicales libres, sino también a su efecto negativo sobre las estructuras de síntesis hormonal de las gónadas y del sistema inmunitario (Nezhdanov et al., 2017).
 MiaBond 360 no solo contiene polifenoles hidrosolubles, sino también liposolubles. Los polifenoles hidrosolubles favorecen la vitamina C y, mientras que, los polifenoles liposolubles fortalecen y estabilizan las membranas celulares. Gracias a los compuestos anteriormente mencionados, podemos aliviar el estrés oxidativo producido por las micotoxinas.
MiaBond 360 no solo contiene polifenoles hidrosolubles, sino también liposolubles. Los polifenoles hidrosolubles favorecen la vitamina C y, mientras que, los polifenoles liposolubles fortalecen y estabilizan las membranas celulares. Gracias a los compuestos anteriormente mencionados, podemos aliviar el estrés oxidativo producido por las micotoxinas.
La zearalenola es una de las micotoxinas más frecuentemente detectadas en los cereales y piensos animales de todo el mundo. Se une a receptores de estrógenos, ejerciendo efectos estrogénicos en varias especies animales.
La microbiota del rumen puede degradar ciertas micotoxinas en sustancias menos tóxicas y ejercer así una función protectora. Sin embargo, en el caso de la zearalenola, la metabolización mediada por la microbiota del rumen podría no resultar en detoxificación.
La zearalenola es convertida principalmente en α-zearalenola y, en menor medida, en β-zearalenola, en el rumen (Gruber-Dorninger et al., 2021). La α-zearalenola es 60 veces más potente que la zearalenola; por el contrario, la β-zearalenola tiene solo 0,2 veces su potencia (Rivera-Chacon et al., 2024).
La zearalenola daña principalmente el sistema reproductor y reduce la fertilidad en el ganado vacuno lechero.
![]() La óptima estructura superficial de la bentonita en MiaBond 360 permite que se ligue eficazmente en rumiantes, evita su adsorción y facilita su excreción a través de las heces.
La óptima estructura superficial de la bentonita en MiaBond 360 permite que se ligue eficazmente en rumiantes, evita su adsorción y facilita su excreción a través de las heces.
El desoxinivalenol (DON) y otros tricotecenos pueden interferir en la síntesis de las proteínas, lo que puede reducir las poblaciones de glóbulos blancos, condicionando y limitando la producción de importantes mediadores inflamatorios (Gott, 2016).
Además, el DON y otros tricotecenos provocan la segregación de la hormona de la saciedad colecistoquinina (CCK), induciendo una sensación de llenado y reduciendo la ingesta de pienso (Artavia, 2024).
 Las fumonisinas (FUM) son micotoxinas que se detectan habitualmente en los silos de maíz. Los microorganismos del rumen permiten que las FUM pasen sin ser metabolizadas en su mayor parte. Al contrario que otras micotoxinas, la metabolización o detoxificación de las FUM en el rumen es escasa (Kemboi et al., 2020).
Las fumonisinas (FUM) son micotoxinas que se detectan habitualmente en los silos de maíz. Los microorganismos del rumen permiten que las FUM pasen sin ser metabolizadas en su mayor parte. Al contrario que otras micotoxinas, la metabolización o detoxificación de las FUM en el rumen es escasa (Kemboi et al., 2020).
Se han añadido diversos microorganismos y enzimas al pienso para degradar algunas de las micotoxinas en metabolitos que resulten menos tóxicos en el tracto gastrointestinal (Haladi, 2024).
 No todos los aglutinadores de toxinas contienen enzimas y bacterias. A fin de proteger el ganado vacuno lechero frente a los efectos adversos de las FUM y DON, se ha incluido en MiaBond 360 la enzima fumonisina esterasa y bacterias Coriobacteriaceae.
No todos los aglutinadores de toxinas contienen enzimas y bacterias. A fin de proteger el ganado vacuno lechero frente a los efectos adversos de las FUM y DON, se ha incluido en MiaBond 360 la enzima fumonisina esterasa y bacterias Coriobacteriaceae.
![]() La enzima fumonisina esterasa convierte las fumonisinas en metabolitos no tóxicos.
La enzima fumonisina esterasa convierte las fumonisinas en metabolitos no tóxicos.
![]() Las bacterias Coriobacteriaceae detoxifican DON mediante su transformación en metabolitos no tóxicos.
Las bacterias Coriobacteriaceae detoxifican DON mediante su transformación en metabolitos no tóxicos.
Bibliografía
Ametaj, B. N., Zebeli, Q., Iqbal, S. (2010). Nutrition, microbiota, and endotoxin-related diseases in dairy cows. R. Bras. Zootec., 39, 433-444.
Andersen, P. H. (2003). Bovine Endotoxicosis-Some aspects of relevance to production diseases. A review. Acta vet. scand., 98, 141-155.
Artavia, I. (2024). Managing mycotoxins for maximum productivity of beef and dairy cattle. Feed and Additive Magazine, March 2024, Year: 4, Issue:38, 51-53.
BonDurant R. H. (2004). Controlling what we can control:Limiting embryonic /fetal losses. The AABP Proceedings, 37, September, 2004, 97-111.
Chodorowska, Z. (2016). Hidden health threats of upsizing. Science & Solutions, a magazine of Biomin, Ruminants, Issue 37, 2016.
Cullor, J. S. (2003). Mastitis and its influence upon reproductive performance in dairy cattle. p.: 176 Proceedings of the international symposium on bovine mastitis
Indianapolis, IN. National Mastitis Council Inc. and American Association of Bovine Practitioners, Arlington, VA.
Dogan, V., Dal, S. D. (2022). Negative effects of zearalenone on reproductive productivity in dairy cattle. Veterinary Journal of Kastamonu University, 1(1), 42-57.
Duchateau, A. (2024). Mycotoxins and endotoxins: Are there any synergisms at stake?
Feed and Additive Magazine, March 2024, Year: 4, Issue:38, 30-34.
Eckel, E. F., Ametaj, B. N. (2016). Invited review: Role of bacterial endotoxins in the etiopathogenesis of periparturient diseases of transition dairy cows. J. Dairy Sci., 99, 5967-5990.
Eckel, E. F., Ametaj, B. N. (2020). Bacterial endotoxins and their role in periparturient diseases of dairy cows: Mucosal vaccine perspectives. Dairy, 1, 61-90.
Garcia, D. D. (2023). Mycotoxin control: Focusing on the problem. The animal nutrition media, nutriNews.
Gott, P. (2016). Mycotoxins, mastitis and milk. Science & Solutions, a magazine of Biomin, Ruminants, Issue 37, 2016.
Graham, M. R. (2014). Effects of lipopolysaccharide induced inflammatory response on early embryo survival in ewes. Master of Science in Reproductive Physiology, Thesis, West Virginia University, USA.
Gruber-Dorninger, C., Faas, J., Doupovec, B., Aleschko, M., Stoiber, C., Höbartner- Guꟗl, A., Schöndorfer, K., Killinger, M., Zebeli, Q., Schatzmayr, D. (2021). Metabolism of zearalenone in the rumen of dairy cows with and without application of a zearalenone-degrading enzyme. Toxins, 13(84).
Haladi, S. (2024). Mycotoxin binder is a misnomer. Feed and Additive Magazine, March 2024, Year: 4, Issue:38, 36-38.
Jackson, S. (2023). Endotoxins in animal feed and their relationship with mycotoxins. NutriNews International, September 2023.
Jiang, Y., Hansen, P. J., Xiao, Y., Amaral, T. F., Vyas, D., Adesogan, A. T. (2019).
Aflatoxin compromises development of the preimplantation bovine embryo through mechanisms independent of reactive oxygen production. J. Dairy Sci., 102, 10506-10513.
Kemboi, D. C., Antonissen, G., Ochieng, P. E., Croubels, S., Okoth, S., Kangethe, E. K., Faas, J., Lindahl, J. F., Gathumbi, J. K. (2020). A review of the impact of mycotoxins on dairy cattle health: Challenges for food safety and dairy production in Sub-Saharan Africa. Toxins, 12(222).
Lavon, Y., Leitner, G., Moallem, U., Klipper, E., Voet, H., Jacoby, S., Glick, G., Meidan, R., Wolfenson, D. (2011). Immediate and carryover effects of Gram-negative and Gram-positive toxin-induced mastitis on follicular function in dairy cows. Theriogenology, 76(5), 942-953.
McCooey, P. (2024). Understanding mycotoxin risk and the impact on egg and chick quality. Feed and Additive Magazine, March 2024, Year: 4, Issue: 38, 26-28.
Melendez, P., Bartolome, J., Archbald, L. F., Donovan, A. (2003). The association between lameness, ovarian cysts and fertility in lactating dairy cows. Theriogenology, 59, 927-937.
Nährer, K. (2015). Mycotoxins and their effects in animals. In: Guide to mycotoxins, Ed.: E. M. Binder, Biomin Edition, Erber AG, Austria, p.: 49-88.
Nezhdanov, A. G., Mikhalev, V. I., Lozovaya, E. G., Lobodin, K. A., Safonov, V. A. (2017).
Pathophysiological aspects of embryonic mortality in dairy cows. Agricultural Biology, 52(2), 338-348.
Nocek, J. E. (1997). Bovine Acidosis: Implications on laminitis. Journal of Dairy Science, 80(5), 1005-1028.
Parker-Norman, S. (2024). Leaky gut and the cost of inflammation. Feed and Additive Magazine, March 2024, Year: 4, Issue:38, 42-43.
Rani, P., Dutt, R., Singh, G., Chandolia, R. K. (2018). Embryonic mortality in cattle-A review. Int. J. Curr. Microbiol. App. Sci., 7(7), 1501-1516.
Rivera-Chacon, R., Hartinger, T., Castillo-Lopez, E., Lang, C., Penagos-Tabares, F., Mühleder, R., Atif, R. M., Faas, J., Zebeli, Q., Ricci, S. (2024). Duration of zearalenone exposure has implications on health parameters of lactating cows. Toxins, 16(116).
Rodrigues, I., Nährer, K., Schwab, C. (2015). Counteracting mycotoxins. In: Guide to mycotoxins, Ed.: E. M. Binder, Biomin Edition, Erber AG, Austria, p.: 141-162.
Seglar, B. (2024). Mycotoxin effects on dairy cattle. [online]. Available from: https://fyi. extension.wisc.edu/forage/mycotoxin-effects-on-dairy-cattle/ [Accessed 14-07-2024].
Szelenyi, Z., Szenci, O., Bodo, S., Kovacs, L. (2023). Noninfectious causes of pregnancy loss at the late embryonic/early fetal stage in dairy cattle. Animals, 13(3390).
Thatcher, W. W., Santos, J. E. P. (2003). Characterization of early embryonic death and prevention of pregnancy wastage. The AABP Proceedings, 36, September 2003, 100-108.
Tournay, A. L. (2024). Navigating mycotoxin challenges in ruminants. Feed and Additive Magazine, March 2024, Year: 4, Issue:38, 39-40.
Walker, S., Smith, R. F., Jones, D. N., Routly, J. E., Morris, M. J., Dobson, H. (2008). The effect of a chronic stressor, lameness, on detailed sexual behaviour and hormonal profiles in milk and plasma of dairy cattle. Reprod. Domest. Anim., 45, 109-117.
Walsh, S. W., Williams, E. J., Evans, A. C. O. (2011). A review of the causes of poor fertility in high milk producing dairy cows. Animal Reproduction Science, 123, 127-138.
Whitetaker, T. B., Slate, A. B. (2015). Sampling and sample preparation for mycotoxin analysis. In: Guide to mycotoxins, Ed.: E. M. Binder, Biomin Edition, Erber AG, Austria, p.: 89-118.
Yildiz, D. (2024). Feed mycotoxin binders and global market status. Feed and Additive Magazine, March 2024, Year: 4, Issue:38, 78-82.
Suscríbete ahora a la revista técnica de nutrición animal
AUTORES

Achaparramiento del maíz: sus efectos sobre la calidad nutricional
Virginia Fain Binda
Harina zootécnica: Ficha de materia prima
Alba Cerisuelo
Efecto de Detoxa® Plus sobre los parámetros productivos en gallinas de postura
Cecilia Rodriguez
Optimizando el desempeño en situaciones de estrés
Horta Felipe
TechnoSpore: La cepa probiótica de Biochem que une lo mejor de dos mundos

El impacto de la suplementación de fibra dietética en lechones
Gustavo Cordero
Grasas oxidadas en porcinos: riesgos y consideraciones. Parte III
María Alejandra Pérez Alvarado
Proteína unicelular: alternativa sostenible a la proteína convencional
Meryem El Kissi
Nutrición de las terneras: Claves para asegurar el éxito. Parte II
José Luis Repetto
Carbohidratos para gatos ¿alternativa energética o enemigo silencioso? Parte II
Ada E. Lugo