


La digestión postruminal comprende los procesos digestivos en el abomaso, intestino delgado y grueso.
En esta etapa se lleva a cabo la absorción de nutrientes provenientes de la fermentación ruminal y de los alimentos no degradados previamente, desempeñando un papel fundamental en el aprovechamiento energético y proteico del animal y además inevitable.
El estudio de la digestión postruminal es esencial para comprender la eficiencia alimentaria y optimizar la nutrición en los sistemas de producción animal, especialmente durante la fase de alta productividad.
Es aquí, en donde se absorben los nutrientes de alto valor biológico, los que son el resultado de la degradación ruminal, la síntesis microbiana y necesariamente los no degradables.
En animales de élite, los nutrientes absorbidos a través de la pared ruminal son insuficientes por sí solos, haciendo que la eficiencia intestinal y el control del paso de nutrientes sean determinantes.
Este proceso fisiológico fundamental se desarrolla en tres etapas claves dentro del tracto posterior del animal:
1.En el abomaso se secreta ácido clorhídrico y enzimas proteolíticas (como la pepsina).
Al disminuir el pH, la acidez desnaturaliza las proteínas que escapan del rumen (proteína no degradable o PND) y la proteína microbiana, preparándolas para ser digeridas de forma eficiente en el intestino.
2.El Intestino delgado es el lugar en donde el animal incorpora los nutrientes al resto del organismo.
La digestión de las proteínas se realiza por medio de las enzimas pancreáticas (tripsina, quimotripsina) y las secreciones intestinales, es aquí en donde se reducen los péptidos a aminoácidos libres, los cuales son absorbidos directamente en la pared intestinal.
La digestión de carbohidratos de reserva, parte del almidón logra pasar la fermentación microbiana en el rumen y es digerido en el intestino delgado por la enzima α-amilasa pancreática y la maltasa intestinal, transformándose en glucosa.
Esta es la vía más eficiente para aportar energía y glucosa al animal.
En el caso de las grasas, las sales biliares emulsionan las grasas y la lipasa pancreática las descompone para su posterior absorción.
3.El intestino grueso actúa, como una cámara de fermentación secundaria.
Los microorganismos que llegan desde el abomaso y el intestino delgado fermentan junto con la parte de la fibra residual, que no se ha degradado en el rumen, produciendo ácidos grasos volátiles (AGV) de cadena corta que se absorben directamente en el colon.
 La digestión postruminal en animales de alta productividad (por ejemplo, vacas lecheras en pico de producción o terneros en cebaderos intensivos) representa un momento crítico para el aporte de proteína metabolizable (PM) y energía neta (EN).
La digestión postruminal en animales de alta productividad (por ejemplo, vacas lecheras en pico de producción o terneros en cebaderos intensivos) representa un momento crítico para el aporte de proteína metabolizable (PM) y energía neta (EN).
A niveles de consumo extremos (>4% del peso vivo), la velocidad de paso de la partícula (kp) se eleva drásticamente (kp ≥0,08h-1 (es el 8% de vaciado por hora)), limitando el tiempo de retención en el retículo-rumen y enviando una gran fracción de sustancias quimo digerible hacia el tracto posterior.
El abomaso funciona como un biorreactor ácido de flujo continuo, optimizado para degradar las estructuras proteicas complejas y la biomasa microbiana mediante la secreción activa de protones (H+) por las células parietales mediante la bomba de protones (H+/K+ -ATPasa) que baja el pH intra abomasal a rangos de 2,0 a 3,0.
Este gradiente rompe los puentes hidrógeno y desnaturaliza las estructuras terciarias y cuaternarias de las proteínas libres.
Por otro lado, provoca la lisis osmótica del flujo de bacterias, arqueas y protozoos ruminales, exponiendo todas sus estructuras a la digestión.
- La activación zimogénica (zimógenos: precursores inactivos de enzimas), el pepsinógeno segregado por las células peptídicas o células principales se cataliza por el pH ácido.
- El pH ácido Acelera la ruptura autocatalítica del pepsinógeno, hasta transformarlo en pepsina, que es una endopeptidasa.
- La endopeptidasa hidroliza preferentemente enlaces en los que intervienen aminoácidos aromáticos (fenilalanina, triptófano, tirosina).
- Ésta fragmenta los polímeros en oligopéptidos de cadena corta.
El quimo ácido pasa al duodeno, activando mecanismos neuroendocrinos (secretina y colecistoquinina) que inducen la secreción biliar y pancreática, neutralizando el pH hasta 6 – 6,5, el óptimo para las enzimas intestinales (Figura 1):
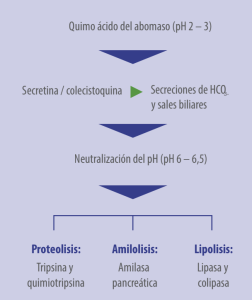
Figura 1. pH y su relación con la actividad enzimática.
El intestino delgado es el responsable de la absorción de aminoácidos, en donde la digestibilidad real de la proteína microbiana ronda del 80-85%.
En la fase luminal la enterocinasa de la mucosa duodenal activa el tripsinógeno a tripsina, esta, a su vez, activa a la quimotripsina, elastasa y carboxipeptidasas A y B, el resultado que se obtiene es la degradación de los oligopéptidos a tripéptidos, dipéptidos y finalmente aminoácidos libres.
En la fase apical o fase de absorción los di y tripéptidos se absorben mediante transporte activo secundario acoplado a un gradiente de protones a través del cotransportador PepT1. Los aminoácidos libres utilizan sistemas transportadores específicos dependientes de sodio.
Una vez dentro del enterocito, las peptidasas citosólicas hidrolizan los péptidos restantes antes de su exportación al sistema porta hepático (Cuadro 1).
En relación a los lípidos que entran al duodeno del rumiante están altamente biohidrogenados por los microorganismos ruminales, presentándose predominantemente como ácidos grasos libres saturados (principalmente ácido esteárico, C18:0) unidos a partículas de la dieta.
El quimo carece de monoglicéridos (sustancias emulsificantes clave) porque los triglicéridos ya fueron hidrolizados en el rumen.
Por lo tanto, los rumiantes dependen críticamente de la lisolecitina (derivada de la fosfatidilcolina biliar tras la acción de la fosfolipasa A2 pancreática) y de las sales biliares para formar micelas mixtas estables a pH ácido.
Siguiendo el tránsito, aproximadamente del 5 al 15% de la materia orgánica total digerible se fermenta en el intestino grueso. Aquí no hay secreción de enzimas digestivas endógenas del hospedador; toda la actividad es microbiana.
| De forma genérica lo que llega al intestino grueso son los residuos de la pared celular vegetal (FND no degradada en rumen), almidón postruminal remanente y proteínas no degradadas y las endógenas (células de descamación y enzimas). |
La microbiota cecal y del colon fermenta estos carbohidratos mediante rutas análogas a las ruminales, produciendo ácidos grasos volátiles (AGV): acetato, propionato y butirato, junto con gases principalmente metano (CH4), y bióxido de carbono (CO2).
Estos AGV se absorben por difusión simple y aportan entre el 5 y el 10% de la energía neta del animal.
El nitrógeno de las proteínas bacterianas sintetizadas de novo en el intestino grueso no puede ser absorbida por el hospedador debido a la ausencia de transportadores de aminoácidos y péptidos.
Todo este nitrógeno se pierde por las heces (nitrógeno metabólico fecal), siendo muy exigentes podríamos decir que representa una ineficiencia neta en el balance de aminoácidos de animales de alta producción.
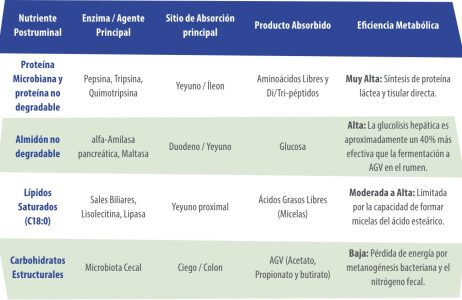
Cuadro 1. Balance y eficiencia nutricional postruminal.
En el cuadro anterior se resumen los coeficientes de digestibilidad aparente y el destino metabólico neto en el tracto posterior al rumen-retículo.
| La formulación avanzada en rumiantes altamente productivos busca un balance termodinámico preciso, para ello se requiere utilizar las ecuaciones de dinámica de flujo o en los coeficientes de absorción específicos por aminoácido bajo los modelos de CNCPS (Universidad de Cornell) o INRAE. |
El flujo de un nutriente (i) que escapa del rumen hacia el resto del tracto se calcula mediante la relación competitiva entre la tasa de degradación ruminal (kd) y la tasa de paso (kp):
 En modelos avanzados (CNCPS), la kp postruminal o tasa de vaciado de los compartimentos posteriores se modeliza de forma diferente:
En modelos avanzados (CNCPS), la kp postruminal o tasa de vaciado de los compartimentos posteriores se modeliza de forma diferente:
Líquidos y partículas ultrafinas, siguen una cinética de primer orden influenciada directamente por el consumo de materia seca en función del peso vivo (% consumo medio diario (CMD) / peso vivo (PV)).
Ecuación de predicción de pasaje para concentrados:
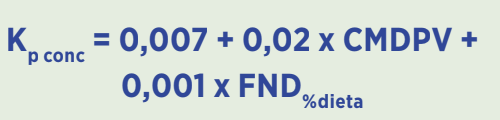 (En donde Kp conc es la tasa de paso del concentrado, CMDPV es el consumo como porcentaje del peso vivo y FND%dieta es el porcentaje de fibra neutro detergente de la dieta).
(En donde Kp conc es la tasa de paso del concentrado, CMDPV es el consumo como porcentaje del peso vivo y FND%dieta es el porcentaje de fibra neutro detergente de la dieta).
Por ejemplo, en vacas de muy alta producción, el cálculo de esta fórmula lleva a la kp por encima de 0,09 h-1, reduciendo drásticamente la digestión ruminal y sobrecargando mecánicamente el intestino delgado.
El CNCPS ha dividido en fracciones la matriz de digestibilidad intestinal específica (Cuadro 2), una vez que el bolo ingresa al duodeno, los modelos no asumen una digestibilidad estática, sino coeficientes de digestibilidad intestinal verdaderos (DIv) asignados a fracciones químicas que no se modifican en su paso a través del rumen:
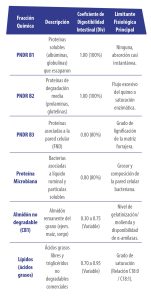
Cuadro 2. Fracciones de digestibilidad intestinal CNCPS.
El objetivo de trabajar con un modelo dinámico de absorción de aminoácidos y el concepto de “Proteína Ideal”, es predecir el flujo de aminoácidos metabolizables (AAM) hacia el yeyuno.
Los sistemas CNCPS e INRAE aplican ecuaciones vectoriales para resolver la absorción neta:
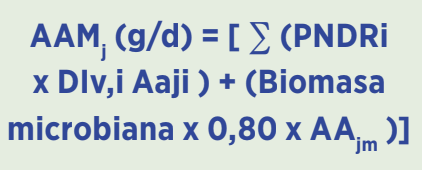 AAMj: Flujo de aminoácidos metabolizables al yeyuno; AAji: Perfil de aminoácidos de la fracción de alimento i para el aminoácido j; AAjm: Perfil de aminoácidos de la biomasa microbiana; EAPj: Eficiencia de absorción después de entrar al enterocito (pérdidas por el metabolismo propio del enterocito antes de llegar a la vena porta).
AAMj: Flujo de aminoácidos metabolizables al yeyuno; AAji: Perfil de aminoácidos de la fracción de alimento i para el aminoácido j; AAjm: Perfil de aminoácidos de la biomasa microbiana; EAPj: Eficiencia de absorción después de entrar al enterocito (pérdidas por el metabolismo propio del enterocito antes de llegar a la vena porta).
La mucosa intestinal consume una gran cantidad de aminoácidos para su propio mantenimiento celular, debido a la alta velocidad de recambio de los enterocitos.
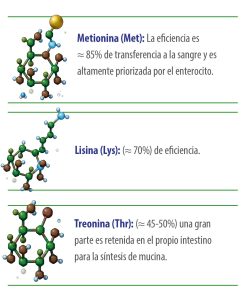
Los aminoácidos esenciales se absorben con eficiencias netas variables hacia el sistema porta:
 El aprovechamiento de los carbohidratos a nivel postruminal presenta ciertas características diferenciales. La digestión de glucosa en el intestino delgado es un 42% más eficiente energéticamente que su fermentación ruminal a propionato (se evitan las pérdidas de metano y calor de fermentación).
El aprovechamiento de los carbohidratos a nivel postruminal presenta ciertas características diferenciales. La digestión de glucosa en el intestino delgado es un 42% más eficiente energéticamente que su fermentación ruminal a propionato (se evitan las pérdidas de metano y calor de fermentación).
Sin embargo, la capacidad del rumiante para secretar α-amilasa pancreática está limitada por la escasez evolutiva de transportadores SGLT-1.
Si el flujo de almidón al duodeno excede los 1,5 a 2 kg/vaca/día, la tasa de absorción celular de glucosa se satura (Vmax alcanzada).
El exceso de almidón genera:
- Aumento de presión osmótica intestinal: Diarreas mecánicas por retención de agua en la luz intestinal.
- Llegada de almidón al ciego en exceso, provocando acidosis cecal (caída del pH del intestino grueso por acumulación de lactato y AGV), lesionando el epitelio del colón y proliferando los microorganismos.
La digestión postruminal de las grasas tiene algunas particularidades: los ácidos grasos que más llegan al intestino son los saturados de cadena larga (como el ácido esteárico, C18:0) que poseen un punto de fusión elevado (≈ 69,3o C). A la temperatura corporal del rumiante (39o C), son sólidos e hidrofóbicos.
Por otro lado, la eficiencia de absorción intestinal de las grasas decae en forma logarítmica a medida que aumenta la dosis total, debido a la saturación de las sales biliares y a la disponibilidad de la lisolecitina para formar las micelas que es relativamente baja.
Si se supera el límite de síntesis de micelas, el ácido esteárico precipita y se excreta intacto por las heces, reduciendo la digestibilidad de la grasa total de la dieta.






